Дифференцировочные торможения
Клеточные аналоги дифференцировочного торможения вырабатывались классическими приемами чередования подкрепляемых и неподкрепляемых стимулов, обычно одной сенсорной модальности, но с различными физическими параметрами.
Как показали Jasper и соавторы (1969), предъявление дифференцировочных стимулов при выработке у обезьян двигательных условных рефлексов на свет вызывало как угнетение активности клеток, так и повышение частоты их разрядов, которое сопутствовало торможению условного двигательного акта. При этом количественные характеристики соотношения нейронов с противоположным знаком реакции на дифференцировочный стимул были различными в разных корковых структурах. В двигательной и сомато-сенсорной корковых зонах 39% клеток реагировали на отрицательный световой раздражитель торможением разрядов, а 61% - активацией; в лобной коре 75% клеток реагировали на дифференцировку торможением активности и 25% - повышением частоты разрядов; в теменной коре дифференцировочный стимул вызывал угнетение активности у 65% нейронов и у 35% клеток - активацию.
Весьма высокий процент клеток двигательной коры, которые реагировали на тормозной стимул повышением частоты разрядов, авторы связывают с активацией мышц-антагонистов условнорефлекторного сгибания конечности. Однако такое объяснение едва ли приемлемо, так как положительные и отрицательные условные реакции клеток возникают задолго до появления поведенческих актов и, как установили Buchwald с соавторами (1961), сопутствующих им условных разрядов системы альфа-мотонейронов, идентифицированных как непременный компонент условнорефлекторного движения.
В этом первом исследовании клеточных аналогов дифференцировочного торможения Jasper с соавторами не соотносили, к сожалению, характер реакции клеток на отрицательный стимул с формой их условной, активности, от знака которой зависит возбуждающее или угнетающее действие тормозного сигнала, как это имеет место при угашении. Такой анализ крайне важен, так как на положительный условный сигнал клетки исследованных образований коры также реагировали не однозначно: в двигательной и сомато-сенсорной коре преобладали нейроны с активирующими реакциями на сигнальный стимул (77% клеток), а в лобной и теменной коре - нейроны с тормозными условными реакциями (62 и 65% клеток соответственно). Между тем даже примерное сопоставление знака реакции клетки на дифференцировочный стимул со знаком предшествующих ответов этой же клетки на положительный сигнал позволило бы оценить степень участия нейронов каждой из корковых структур в становлении внутреннего торможения как системного процесса.
В последующих работах дифференцировочное торможение изучалось на нейронах ряда корковых образований мозга при выработке различного рода условных рефлексов, классических и инструментальных (Morrell, 1960; Evarts, 1966; Travis, Sparks, 1968; Hirano e. a., 1970; Engel, Woody, 1972, и др.). Нейроны с активирующими условными реакциями, которые обычно испытывались на дифференцировки, на отрицательный стимул либо не реагировали, либо отвечали торможением активности. Так, Travis и Sparks (1968) обнаружили, что при выработке у обезьян пищедобывательных условных рефлексов большинство нейронов бледного шара из числа проявивших способность дифференцировать подкрепляемые и неподкрепляемые звуковые стимулы реагировало повышением частоты разрядов на положительный сигнальный стимул и не отвечало на отрицательный; только некоторые клетки бледного шара в отдельных пробах отвечали на дифференцировочный стимул снижением частоты разрядов.
У. Г. Гасанову (19726) удалось, однако, зарегистрировать активные тормозные реакции нейронов слуховой коры на дифференцировочные стимулы при выработке у кошек (с хронически вживленными микроэлектродами) условных мигательных рефлексов на звук. По мере упрочения дифференцировочного торможения клетки с активирующими условными реакциями отвечали на неподкрепляемые звуковые стимулы снижением частоты разрядов, которое обычно совпадало с отсутствием условнорефлекторного мигания. Интересно, что у некоторых нейронов выработка дифференцировочного торможения оказывала влияние на реализацию положительной условной реакции, которая также приобретала тормозные черты в виде некоторого снижения частоты разрядов, т. е. имела место, по-видимому, своеобразная генерализация тормозного процесса в пределах нейрона.
Детальный анализ развития дифференцировочного торможения у нейронов различных корковых и подкорковых образований мозга при выработке у крыс условных пищедобывательных рефлексов на звук провели Olds с сотрудниками (Hirano, Best, Olds, 1970; Segal, Olds, 1972; "Disterhoft, Olds, 1972). Весьма важной, с нашей точки зрения, находкой Hirano, Best и Olds (1970) оказалась приуроченность дифференцировочного торможения в виде отсутствия реакции на отрицательный стимул к моменту проявления предшествующих условных реакций клетки на положительный сигнал с латентными периодами, превышающими 300 мс, т. е. тормозились только так называемые стабилизированные условные ответы. Каких-либо различий в реакциях на положительные и отрицательные звуковые стимулы в пределах начальных 300 мс изолированного односекундного действия сигнального стимула не обнаружено.
Эти наблюдения дают дополнительное основание относить к подлинным условнорефлекторным реакциям клетки только ответы, латентные периоды которых значительно возрастают по мере выработки и упрочения условного рефлекса (см. главу III, с. 91). Такого рода реакции на подкрепляемые и неподкрепляемые звучшвые стимулы проявили нейроны гиппокампа, латерального гипоталамуса и ретикулярных структур среднего мозга. Следует отметить также, что нейроны гиппокампа (поля СА-1 и СА-3) проявили статистически достоверную дифференцировку двух звуковых стимулов в пределах первых 25 - 75 предъявлений отрицательного раздражителя, тогда как у клеток ретикулярной формации среднего мозга дифференцировки появлялись после 150 проб. Следовательно, нейроны гиппокампа более чем в 2 раза быстрее "обучались" различать подкрепляемый и не-подкрепляемый раздражители (Hirano e. a., 1970).
Сравнительную оценку способности нейронов различных образований мозга дифференцировать подкрепляемые и неподкрепляемые звуковые стимулы в процессе реализации у крыс пищедобывательных условных рефлексов провели в последующих работах Segal и Olds (1972), Disterhoft и Olds (1972), Segal и Olds на статистически достоверном материале показали, что клетки полей СА-1 и СА-3 гиппокампа значительно более точно дифференцируют оба стимула, чем клетки зубчатой извилины и субикулума. По данным Disterhoft и Olds, нейроны таламической группы ядер, включая внутреннее коленчатое тело, лучше дифференцируют подкрепляемые и неподкрепляемые звуковые стимулы, чем корковые клетки, хотя эти различия не столь существенны. В пределах таламических ядер наиболее достоверные и прочные дифференцировки проявили нейроны заднего ядра таламуса, клетки которого отличались как отчетливой выраженностью условных реакций, так и относительно небольшим числом сочетаний, необходимых для их появления (Disterhoft Olds, 1972).
По данным Phillips (1971), полученным при выработке у крыс пищедобывательных и оборонительных условных рефлексов в хронических экспериментах по Олдсу, способность дифференцировать подкрепляемые и неподкрепляемые звуковые стимулы проявило 80% нейронов внутреннего коленчатого тела и 60% клеток ретикулярной формации среднего мозга. Клетки нижних бугорков, латерального гипоталамуса и дорсальных ядер таламуса не были способны различать эти стимулы. Следует отметить, что краткость сообщения не позволяет оценить приемы идентификации условных связей и дифференцировок, а также степень достоверности полученных результатов.
Masland с соавторами (1972) не удалось выработать дифференцировки на неподкрепляемые световые стимулы у нейронов наружного коленчатого тела при реализации у бодрствующих кошек пищедобывательных условных рефлексов на свет. Все исследованные нейроны (65 клеток) реагировали на оба стимула (вспышки света различной частоты) ответами, различия которых не выходили за пределы вариабельности реакций как на положительный, так и на отрицательный стимул. Авторы допускают, что приемы выработки дифференцировок и анализа результатов могли быть неадекватными для нейронов наружного коленчатого тела, которые и без сочетаний стабильно реагировали на световые стимулы. Допускается также, что дифференцированные реакции клеток этого таламического образования на подкрепляемые и неподкрепляемые световые стимулы могли проявить себя в более тонких различиях временной последовательности разрядов, чем это позволяли выявить постстимульные гистограммы, либо в различной структуре рецептивных полей.
Интересные факты, имеющие отношение к особенностям условной связи и дифференцировочного торможения, описаны Woody и Yarowsky (1972). У бодрствующих кошек вырабатывались мигательные условные рефлексы на подпороговую электростимуляцию корковой зоны проекции орбикулярной мышцы глаза в сочетаниях с постукиванием по переносице, которое вызывало мигание и служило подкрепляющим раздражителем. Контрольная пороговая стимуляция этой же точки коры вызывала мигательный рефлекс с латентными периодами, не превышающими в среднем 8 мс (по данным ЭМГ орбикулярной мышцы). После нескольких сотен сочетаний у животного появлялись условные мигательные рефлексы на ранее неэффективную подпороговую электростимуляцию с латентными периодами в пределах 12 мс. Стимуляция сверхпороговыми токами также вызывала мигательный рефлекс, но латентность его при этом не превышала контрольную величину в 8 мс.
Наблюдение Woody и Yarowsky (1972) может означать, что интенсивность тока в такой форме эксперимента приобретает специфические черты сигнального стимула и уменьшение латентных периодов поведенческого акта до контрольного уровня при стимуляции сверхпороговым током следует рассматривать как дифференцировку на интенсивность стимула. Следовательно, повышение по мере сочетаний латентных периодов вырабатываемой поведенческой реакции представляет собой, как нами неоднократно подчеркивалось, неотъемлемую особенность условного рефлекса, природу и механизмы которой еще предстоит раскрыть. Другие формы поведенческих реакций этими особенностями, по-видимому, не обладают, что может быть использовано как прием идентификации условных связей и дифференцировок в тех экспериментальных ситуациях, когда оба стимула в равной мере наделены способностью вызывать безусловные рефлексы.
В нашей лаборатории дифференцировочное торможение исследовалось на нейронах двигательной и слуховой коры в процессе выработки у бодрствующих кроликов двигательных условных рефлексов сочетаниями звуковых стимулов с электрокожным раздражением передней конечности (Е. Л. Полонская, М. Я. Рабинович, 1971; Rabinovich, Polonskaya, 1973).
Дифференцировки вырабатывались параллельно с положительными условными реакциями на одном и том же нейроне чередованием подкрепляемых и неподкрепляемых звуковых стимулов. После 5-10 предъявлений дифференцировочного стимула примерно 60% клеток из числа испытанных на дифференцировку были способны различать положительные и отрицательные раздражители одной сенсорной модальности, но с различными физическими параметрами. В двигательной коре 18 из 29 клеток, а в слуховой коре 6 из 10 клеток отчетливо дифференцировали подкрепляемые и неподкрепляемые стимулы.
Как в процессе угашения клеточных условных реакций, так и при дифференцировках отрицательный стимул не только перестает вызывать условную реакцию нейрона, но и способен активно ее тормозить. При дифференцировках активирующих условных реакций неподкрепляемый стимул вызывает снижение частоты разрядов нейрона до полного прекращения импульсации. При дифференцировках тормозных условных реакций клетка достаточно часто отвечает на отрицательный стимул повышением частоты разрядов, т. е. имеет место растормаживание дифференцировочным стимулом некоторой популяции нейронов. Примеры торможения и растормаживания дифференцировочным раздражителем условной активности клеток двигательной коры представлены на рис. 27 и 28, клеток слуховой коры - на рис. 31 и 32.
Усредненные на ЭВМ результаты дифференцировок по всем пробам для 15 нейронов двигательной коры (см. рис. 58) и для 6 нейронов слуховой коры (см. рис. 59) показали, что хорошо выраженное снижение частоты разрядов у клеток с активирующими условными реакциями наступает при первых пяти предъявлениях дифференцировочного стимула. При последующих пробах на дифференцировку торможение условной активности значительно усиливается, особенно у нейронов слуховой коры, и частота импульсации оказывается ниже уровня фоновых разрядов до сочетаний. Подобным же образом у нейронов с тормозными условными реакциями эффекты растормаживания, вызванные дифференцировочным стимулом, четко проявляются в первых пяти пробах на дифференцировку и в последующем достигают заметной величины, превышающей уровень фоновой активности до сочетаний.
Как правило, при первых применениях дифференцировочного стимула клетки на него не реагировали, сохраняя при этом ответы на условный раздражитель. Затем наступала кратковременная фаза (1-2 пробы), когда нейрон реагировал на оба стимула - положительный и отрицательный, после чего относительно быстро развивалось хорошо выраженное и достаточно устойчивое угнетение условной активности клеток дифференцировочным стимулом в форме торможения активирующих и растормаживания тормозных условных реакций.
Часть нейронов, у которых не удалось выработать дифференцировок, сохраняла реакции на оба стимула, причем ответы на дифференцировочный раздражитель иногда усиливались по мере его повторения и были хорошо выражены даже тогда, когда положительные условные реакции нейрона падали ниже уровня, принятого за значимый, или вовсе отсутствовали.
В контрольных пробах у 11 нейронов двигательной коры были испытаны активирующие и тормозные реакции па звуковые стимулы тех же параметров, которые чаще всего применялись в качестве условного и дифференцировочного раздражителей (щелчки с частотой 20 и 10 в 1 с). Как и при выработке положительных и отрицательных условных связей, стимулы предъявлялись поочередно до 30-40 раз с варьирующими интервалами в пределах 40-90 с, но ни один из них не подкреплялся. Контрольные опыты показали, что все испытанные клетки реагировали на оба неподкрепляемых стимула примерно равным образом во всех пробах, не дифференцируя их (рис. 60).
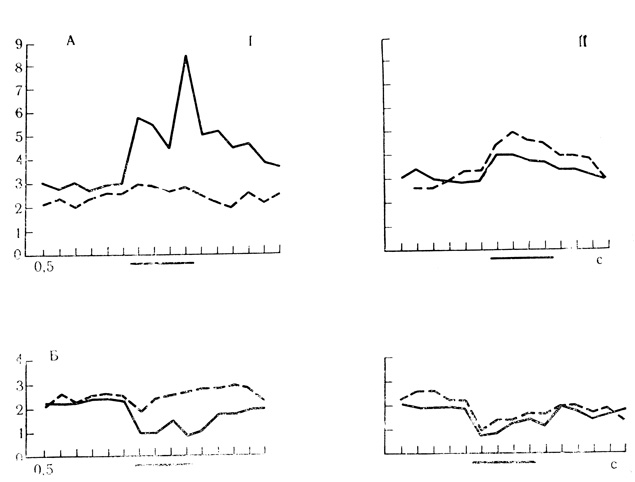
Рис. 60. Контрольные пробы на действие подкрепляемых и неподкрепляемых звуковых стимулов. Нейроны двигательной коры бодрствующих кроликов. А - активирующие реакции клеток: I - реакции на щелчки 20 в 1 с (сплошная линия), подкрепляемые электрокожным раздражением конечности (усреднено 150 сочетаний для 7 клеток) и щелчки 10 в 1 с (прерывистая линия), предъявляемые как дифференцировочный стимул (усреднены 93 пробы для 7 клеток); II - реакции на неподкрепляемые щелчки 20 в 1 с (сплошная линия) и 10 в 1 с (прерывистая линия) в контрольных предъявлениях; для 7 клеток усреднены 104 и 32 пробы соответственно; Б - тормозные реакции клеток: I - реакции на щелчки 20 в 1 с (сплошная линия), подкрепляемые ЭКР конечности (усреднено 250 сочетаний для 8 клеток) и щелчки 10 в 1 с (прерывистая линия), предъявляемые как дифференцировочный стимул (усреднено 113 проб для 8 клеток); II - реакции на неподкрепляемые щелчки 20 в 1 с (сплошная линия) и 10 в 1 с (прерывистая линия) в контрольных предъявлениях; для 4 клеток усреднены 99 и 35 проб соответственно. На оси ординат - число импульсов за 0,5 с (Е. Л. Полонская, 1974)
Представляют интерес также значительные различия в интенсивности спонтанных и вызванных реакций клеток при действии неподкрепляемых (дифференцировочных) стимулов, когда их предъявление чередуется с сочетаниями, и в ситуации, когда те же стимулы предъявляются вне связи с предшествующими сочетаниями.
Как следует из усредненных графиков, представленных на рис. 60, исходный уровень фоновой активности клеток при реализации активирующих условных реакций заметно выше уровня спонтанных разрядов этих же клеток при выработке дифференцировок, тогда как в контрольных пробах исходная спонтанная активность не отличается от фоновых разрядов, которые предшествуют активирующей условной реакции (рис. 60, А, I и II). Это означает, что неподкрепляемый стимул дифференцировочного значения способен снижать возбудимость клетки в межсигнальных интервалах, чего лишены те же сенсорные раздражители, действующие вне предшествующих сочетаний. С другой стороны, в контрольных пробах нейроны реагируют на звуковые щелчки с частотой разрядов, почти в 2 раза превышающей интенсивность реакции на те же стимулы, когда они предъявляются как дифференцировочные после предшествующего и активирующего клетку сочетания (см. рис. 60, А, I и II).
Те же соотношения имеют место у клеток с тормозными условными реакциями: растормаживание ответов на дифференцировочный стимул и отсутствие растормаживающего действия того же стимула в контрольных пробах (см. рис. 60, Б, I и II).
Столь заметные различия в характере фоновой и вызванной активности клеток в различных условиях предъявления одних и тех же стимулов могут служить указанием на возможные механизмы активного воздействия дифференцировочного торможения, которое связано, по-видимому, с влиянием на клетку подкрепляющего фактора в предшествующем дифференцировке сочетании. В пользу такого предположения косвенно свидетельствуют известные факты повышения уровня фоновой активности нейрона при выработке активирующих условных реакций и снижения - при выработке тормозных реакций (Н. Н. Василевский, 1968; В. Б. Швырков, 1968; Б. И. Котляр, 1970; O'Brien, Fox, 19696; Ben-Ari, 1972; Weinberger e. a., 1972, и др.).
Сопоставляя условия предъявления дифференцировочных стимулов, которые всегда чередуются с сочетаниями, с условиями предъявления тех же раздражителей в контрольных пробах, можно предположить, что активная природа дифференцировочного торможения поддерживается механизмами типа неспецифической гетеросинаптической сенситизации. Такого рода механизмы складываются в результате воздействий на клетки подкрепляющего фактора в каждом из предшествующих сочетаний и реализуются последующим действием дифференцировочного стимула. Наличие механизмов посттетанического гетеросинаптического облегчения реакций нейронов корковых структур, в частности двигательной коры, было показано Л. Л. Ворониным (1970 в, 1971 а).
Можно допустить, следовательно, что клеточные аналоги дифференцировочного торможения реализуются механизмами, которые существуют на нейронах рядом с механизмами выработки условной связи и взаимодействуют по мере чередования подкрепляемых и неподкрепляемых стимулов. Сходные предположения о существовании раздельных механизмов для габитуации и дисгабитуации полисинаптических спинальных рефлексов были ранее высказаны Thompson с соавторами (1972).
Если развитие угасательного торможения на нейроне выступает как процесс размыкания временной связи между двумя видами деятельности в пределах, по-видимому, одних и тех же синаптических входов для общего пути сложившейся ассоциации, то дифференцировка подкрепляемых и неподкрепляемых стимулов опосредуется, должно быть, разными синаптическими входами для этих раздражителей через соответствующие системы вставочных нейронов активирующего и тормозного действия. В зависимости от того, какого типа интернейроны включены в полисинаптические пути условной связи, дифференцировочное торможение будет проявляться или как угнетение активирующих условных реакций клетки, или как растормаживание клеточных реакций с тормозной условной активностью.
Вероятно, только потому, что два такого рода механизма сосуществуют на одной и той же клетке, последняя располагает способностью адекватно реагировать на подкрепляемые и неподкрепляемые стимулы одной сенсорной модальности ответами с противоположным знаком активности.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'