Угасательное торможение
Угашение условных реакций отменой подкрепляющего стимула оказалось наиболее исследованным аналогом внутреннего торможения на клеточном уровне. Вероятно, это связано как с исторически сложившейся традицией в лабораториях И. П. Павлова, так и с относительно простой процедурой выработки этой формы торможения.
У большинства нейронов различных корковых и подкорковых образований мозга угашение условных реакций наступает сравнительно быстро (в пределах до 30 предъявлений неподкрепляемого сигнального стимула) независимо от приемов выработки условных связей - модельных, классических или инструментальных. При этом чем более упрочена условная реакция клетки предшествующими сочетаниями, тем больше требуется предъявлений неподкрепляемого раздражителя для ее угашения (В. В. Шульговский, Б. И. Котляр, 1968; Л. Г. Воронин, 1969; Б. И. Котляр, 1970). Восстановление угашенных условных реакций также зависит от предшествующего числа сочетаний: чем более упрочена условная связь, тем быстрее она освобождается от тормозного воздействия, что связано, как предполагают, со степенью консолидации следовых процессов (Л. Г. Воронин, 1969; O'Brien, Fox, 1969a). Ряд авторов отметил, что по мере угашения как активирующих, так и тормозных условных реакций наиболее устойчивым оказывается следовой компонент подкрепления, т. е. та фаза условной реакции клетки, которая при сочетаниях совпадала с действием подкрепляющего стимула (Л. Л. Воронин. Р. Г. Кожедуб, 19716; Н. Н. Василевский и др., 1972а; Kamikawa e. a., 1964).
Действие угашающего стимула проявляется в более или менее выраженном торможении условной активности нейрона, которое постепенно развивается по мере повторных предъявлений неподкрепляемого сигнала. У клеток с активирующими условными реакциями угашение обычно описывается как снижение частоты условных разрядов до исходного уровня фоновой активности (Б. И. Котляр, 1969; Yoshii, Ogura, 1960; Evarts, 1966; Fetz, 1969; Fuster, Uyeda, 1971, и др.). Вместе с тем отрицательный стимул не только перестает вызывать условные реакции, но и способен активно тормозить фоновую активность клетки до полного прекращения импульсации (Н. Н. Василевский, 1968; М. Я. Рабинович, 1969; Л. Л. Воронин, Р. Г. Кожедуб, 1971а; Kamikawa e. a., 1964).
Клетки с тормозной условной активностью при угашении часто реагируют на действие отрицательного стимула повышением частоты разрядов (И. Н. Кондратьева и др., 1970; М. Я. Рабинович, 1970; Г. И. Шульгина, 1971; Kamikawa e. a., 1964; Я. Хори и др., 1967). В этом случае имеет место растормаживание заведомо отрицательными стимулами значительной популяции нейронов, причастных к выработке условного рефлекса. При этом ответ на тормозной стимул в отдельных пробах заметно превышает реакцию клетки на его действие в контрольных предъявлениях до сочетаний.
Растормаживающее действие отрицательного раздражителя при угашении клеточных тормозных условных реакций впервые было зарегистрировано Kamikawa с соавторами (1964) на нейронах некоторых ядер неспецифической проекционной системы таламуса и рассматривалось ими как явление посттормозной отдачи условно-рефлекторной природы. Пользуясь возможностью длительно регистрировать активность нейронов на обездвиженных флакседилом кошках, авторы отметили, что эффекты растормаживания наиболее отчетливо проявлялись при достаточно упроченном внутреннем торможении, т. е. при хорошо выраженных и стабильных тормозных реакциях клеток активирующего типа. Однако при длительном угашении тормозных условных реакций растормаживающее действие отрицательного стимула постепенно шло на убыль и клетки начинали проявлять явную тенденцию к адекватным тормозным реакциям на угашающий раздражитель, углубляя общий процесс внутреннего торможения. Эти важные факты нуждаются в специальном анализе на клеточном и синаптическом уровне.
Представляют интерес данные о некоторых особенностях процесса угашения клеточных условных реакций. К ним относится наблюдение Н. Н. Василевского (1968). Автор обнаружил, что максимальное снижение частоты разрядов нейрона в ответ на отрицательный стимул приходится на период его действия, который соответствовал максимальной условной активности в сочетаниях, т. е. торможение возникает примерно с теми же латентными периодами, которыми, по данным Н. Н. Василевского, характеризуются подлинные условные реакции на клеточном уровне. Это наблюдение иллюстрирует известный вывод И. П. Павлова, что прежде всего тормозятся процессы, больше других вовлеченные в активную деятельность.
В качестве другой особенности следует отметить данные O'Brien и Fox (1969a) о том, что при прочих равных условиях выработки и угашения условного рефлекса тормозные условные реакции клеток угашаются значительно быстрее, чем активирующие. Если для угашения активирующей реакции нейрона требовалось 15-30 предъявлений неподкрепляемого сигнала, то тормозные реакции могли быть полностью угашены в пределах первых пяти проб на угашение. В этом факте авторы усматривают проявление одного из механизмов поддержания стабильности выработанного навыка на достаточно высоком уровне и достаточно длительно даже в процессе развивающегося угасательного торможения.
Существенные различия в развитии угасательного торможения у нейронов некоторых образований лимбической системы (гиппокамп, латеральная зона гипоталамуса, преоптическая область) и ретикулярной формации среднего мозга были обнаружены Hirano с соавторами (1970) при выработке у крыс пищедобывательных условных рефлексов на звуковой стимул. Условные реакции нейронов ретикулярных структур сравнительно легко угашались повторными предъявлениями неподкрепляемого сигнала, частота разрядов постепенно снижалась до контрольного уровня фоновой активности. В противоположность этому клетки гиппокампа (поля СА-1 и СА-3) проявляли при угашении только относительное снижение частоты разрядов по сравнению с действием положительного сигнала. Полное угашение условной реакции было невозможным по крайней мере в пределах 150 предъявлений неподкрепляемого стимула. Нейроны латерального гипоталамуса и преоптическон области занимали промежуточное положение: снижение частоты импульсации в ответ на отрицательный стимул было более выраженным, чем в клетках гиппокампа, но полного угашения условной реакции достичь не удалось.
Особенности угашения условных реакций клеток полей СА-1 и СА-3 гиппокампа Hirano и соавторы (1970) связывают с ролью этих структур в сохранении следовых процессов (энграммы поведенческого акта), которые остаются нетронутыми, когда поведенческий акт угашен. Это может означать, что при угашении условного рефлекса тормозятся преимущественно механизмы воспроизведения следовых процессов.
По нашим данным, условные реакции большинства нейронов двигательной и слуховой корковых зон, а также внутреннего коленчатого тела, выработанные у бодрствующих кроликов сочетаниями звуковых стимулов с электрокожным раздражением конечности, могли быть угашены отменой подкрепляющего стимула и вновь восстановлены повторными сочетаниями. Однако относительное число нейронов, проявивших способность к выработке угасательного торможения, оказалось не одинаковым в различных структурах мозга.
В двигательной коре условные реакции были сравнительно легко угашены у 19 из 27 нейронов, испытанных на угашение (70%). У 8 нейронов условная активность не могла быть угашена, несмотря на относительно большое число проб. В их числе было 7 клеток с неугашенными условными ответами активирующей формы и одна клетка с тормозной условной активностью.
При угашении активирующих условных реакций не-подкрепляемый стимул вызывал заметное снижение частоты разрядов клетки, которое в отдельных пробах доходило до полного прекращения разрядов. Иногда имело место кратковременное восстановление условной активности по ходу угашения, и тогда процесс развития внутреннего торможения выглядел волнообразным. При возобновлении подкрепления условная активность быстро восстанавливалась и достигала значимого уровня после пяти сочетаний (рис. 53).
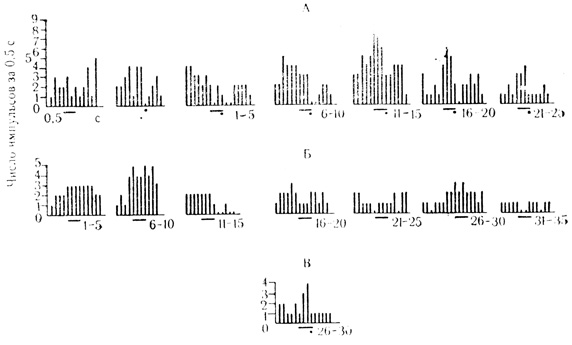
Рис. 53. Гистограммы угашения (Б) и восстановления (В) активирующей условной реакции (А) нейрона двигательной коры. Клетка на глубине 1700 мкм. Под каждой гистограммой - число сочетаний или проб на угашение, которым предшествовало 90 сочетаний на животном. Первые две гистограммы на А - контрольные пробы на звуковой стимул (черта) и электрокожное раздражение (точка). Принцип построения гистограмм тот же, что на рис. 17 (М. Я. Рабинович, Е. Л. Полонская, 1973)
При угашении тормозных условных реакций клетки часто отвечали на отрицательный стимул повышением частоты разрядов, превышающим контрольный уровень фоновой активности, т. е. имело место растормаживающее действие тормозного раздражителя (рис. 54). Растормаживание тормозных условных реакций едва ли играет существенную роль в организации внутреннего торможения поведенческого акта вследствие кратковременного и неустойчивого действия. Однако оно может иметь значение как фактор, умеряющий тормозное состояние на клеточном уровне в процессе формирования внутреннего торможения популяции нейронов, реализующих приспособительный поведенческий акт.
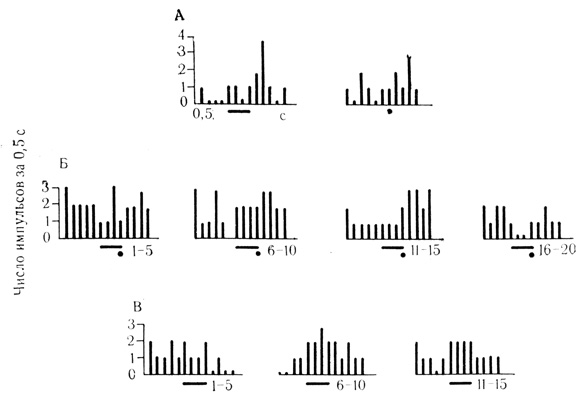
Рис. 54. Гистограммы угашения (В) тормозной условной реакции (Б) нейрона двигательной коры. Клетка на глубине 2000 мкм. Под каждой гистограммой - число сочетаний или проб на угашение, которым предшествовало 119 сочетаний на животном. Первые две гистограммы (А) - контрольные пробы на звуковой стимул (черта) и электрокожное раздражение конечности (точка) (М. Я. Рабинович, Е. Л. Полонская, 1973)
Угашение активирующих условных реакций нейронов двигательной коры наступало в среднем после 10 предъявлений неподкрепляемого раздражителя, а для угашения тормозных условных ответов требовалось еще меньше проб. Как видно на рис. 55, представляющем усредненные на ЭВМ результаты по всем пробам для 18 клеток, у нейронов с активирующими условными реакциями первые 10 предъявлений отрицательного стимула привели только к некоторому снижению частоты разрядов, а полное угашение условной реакции наступало при последующих пробах. Нейроны с тормозными условными реакциями уже в первых 10 пробах на угашение проявили заметное угнетение этой формы условной активности в виде растормаживания реакции с повышением частоты разрядов клеток, которое усиливалось при последующих предъявлениях отрицательного стимула (см. рис. 55, II и III).
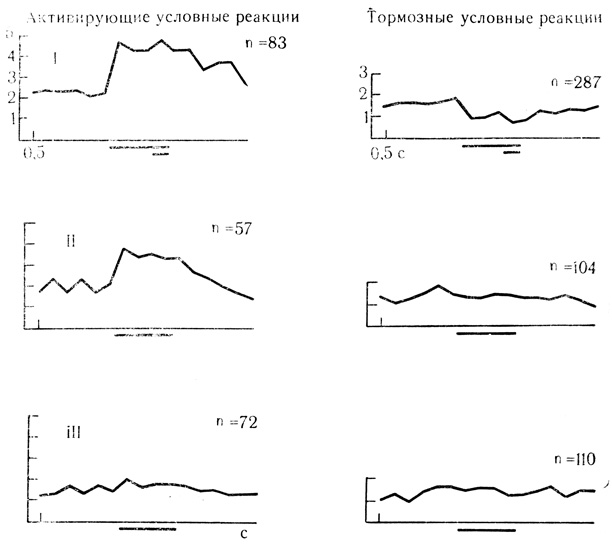
Рис. 55. Динамика выработки угасательного торможения у нейронов двигательной коры с активирующими и тормозными условными реакциями. Активирующие условные реакции: I - усредненные результаты 83 сочетаний (п) для 7 клеток; II - первые 10 проб на угашение для 7 клеток; III - последующие 20 проб на угашение для 5 клеток. Тормозные условные реакции: I - усредненные результаты 287 сочетаний для И клеток; II - первые 10 проб на угашение для 11 клеток; III - последующие 20 проб на угашение для 10 клеток. На оси ординат - число импульсов за 0,5 с. (Е. Л. Полонская, 1974)
В слуховой коре из 12 нейронов, испытанных на угашение, способность к выработке угасательного торможения проявило 6 клеток, у которых после 5-15 предъявлений неподкрепляемого стимула условные реакции исчезали. У других 6 нейронов условная активность не могла быть угашена, несмотря на большое число проб. Как и в двигательной коре, отрицательный стимул активно тормозил условные реакции клеток, что выражалось в снижении частоты разрядов у нейронов с активирующими условными ответами и в некотором повышении частоты разрядов у клеток с тормозной условной активностью (эффекты растормаживания реакции). Угашенные условные реакции легко восстанавливались несколькими повторными сочетаниями. Обращает на себя внимание то, что разряды клетки, приуроченные к моменту действия отмененного подкрепления, часто сохранялись дольше, чем условная активность, и требовалось больше проб, чтобы эта следовая реакция полностью угасла. Примеры развития угасательного торможения у нейронов слуховой коры представлены па рис. 56 и 20.
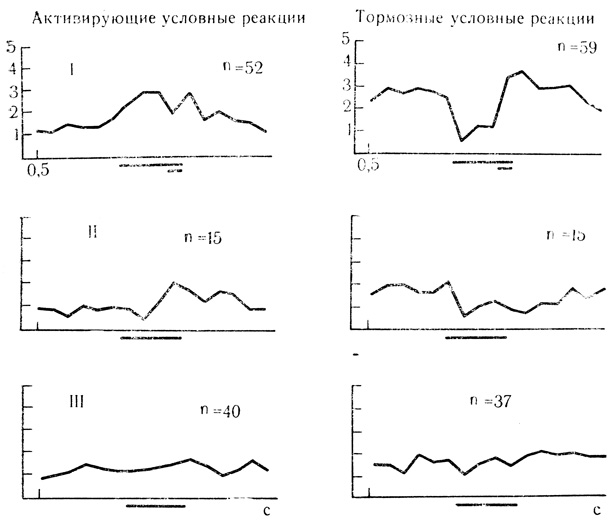
Рис. 56. Выработка угасательного торможения у нейронов слуховой коры с активирующими и тормозными условными реакциями. Активирующие условные реакции: I - усредненные результаты 52 сочетаний для 3 клеток; II - первые 5 проб на угашение для 3 клеток; III - последующие пробы на угашение для 3 клеток. Тормозные условные реакции: I - усредненные результаты 59 сочетаний для 3 клеток; II - первые 5 проб на угашение для 3 клеток; последующие пробы на угашение для 3 клеток (Е. Л. Полонская, 1974)
Особый интерес представляет развитие угасательного торможения (у нейронов внутреннего коленчатого тела (ВКТ), поскольку условные связи у большей части клеток (15 из 27) этого релейного таламического образования были идентифицированы по появлению ответов на ранее недеятельное подкрепление (электрокожное раздражение конечности) и воспроизведению следовой реакции при пропусках его в отдельных сочетаниях с последующим угашением этой реакции. Такой прием идентификации позволил выявить подлинные условные связи у 33% нейронов ВКТ, несмотря на то что в предварительных контрольных пробах почти все клетки этой структуры отвечали на звуковые стимулы (щелчки) стабильными реакциями (см. главу IV, с. 137).
Угашению были подвергнуты условные реакции у 11 нейронов ВКТ. Три клетки из этого числа (две - с активирующими реакциями) относились к нейронам (12 клеток), у которых в результате сочетаний появились ответы на звуковой стимул или изменился знак реакции в соответствии со знаком действия подкрепляющего стимула. После 5-15 предъявлений неподкрепляемого раздражителя условные реакции у этих нейронов были угашены, но могли быть легко восстановлены повторными сочетаниями. При угашении тормозной условной реакции, выработанной в соответствии с угнетающим действием на клетку подкрепления, имело место восстановление характерного начального разряда на звуковой стимул, как это видно на рис. 35.
Восемь клеток ВКТ относились к нейронам (15 клеток), условная связь которых идентифицировалась по появлению активирующей реакции на подкрепление и воспроизведению этой реакции как следового явления при пропусках подкрепления в сочетании. Условные реакции, выработанные по такому типу временной связи, требовали больше проб для угашения и у 6 клеток угасли после 20-30 предъявлений неподкрепляемого стимула. У 2 нейронов условная связь в пределах этого числа проб не могла быть угашена, хотя вероятность появления следовой реакции на подкрепляющий стимул снизилась более чем в 2 раза. Пример угашения следовых разрядов на отмененное подкрепление у отдельного нейрона представлен на рис. 34. Видно, что наряду с постепенным исчезновением следовой реакции на подкрепление заметно уменьшается число импульсов в разряде на сигнальный стимул, достигая уровня контрольных проб.
Динамика угашения следовых разрядов на подкрепляющий стимул, усредненных по всем пробам для 6 нейронов, представлена на графиках рис. 57. В результате сочетаний звуковых щелчков с электрокожным раздражением конечности реакции этих клеток на условный и безусловный стимулы стали сходными по частоте разрядов и отчетливо разделены периодом сниженной активности, который указывает на известное, сходство и структуры ответов (см. рис. 57, I). По мере угашения условной связи следовая активирующая реакция на подкрепление постепенно снижается до контрольного уровня фоновых разрядов. Усредненная реакция на звуковой стимул остается без изменений (см. рис. 57, II и III).
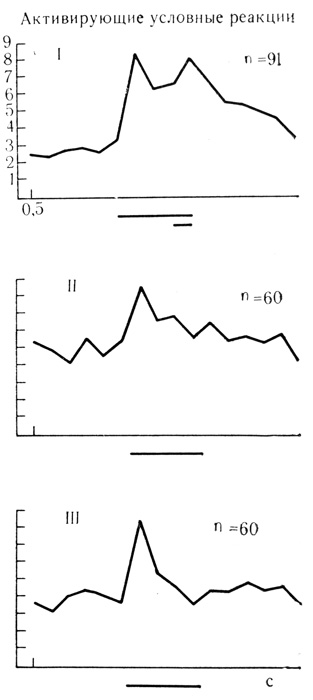
Рис. 57. Выработка угасатель-ного торможения у нейронов внутреннего коленчатого тела, активирующие условные связи которых идентифицировались по появлению и угашению ответов на подкрепляющий стимул. I - усредненные результаты 91 сочетания для 6 клеток; II - первые 10 проб на угашение для 6 клеток; III - последующие пробы на угашение для 6 клеток (Е. Л. Полонскап, 1974)
Пока еще трудно представить, какие механизмы регулируют противоположный характер реакций на подкрепляемые и неподкрепляемые стимулы и где эти механизмы локализуются - в синаптическом аппарате самой клетки или на пути к ней, в многочисленных вставочных нейронах. Л. Л. Воронин и Р. Г. Кожедуб (19716) обнаружили, что при угашении условных ВПСП нейронов моторной коры кролика, выработанных сочетаниями неэффективной и эффективной электростимуляции различных точек корковой поверхности, амплитуда их постепенно снижалась до исходного уровня "синаптического шума" без заметного перехода в гиперполяризацию клеточной мембраны.
Отсутствие ожидаемой гиперполяризации мембраны клетки при развитии угасательного торможения в сложных полисинаптических путях такого рода условной связи допустимо объяснить тем обстоятельством, что наряду с повышением тормозного синаптического притока в какой-то мере сохраняется и возбуждающий синаптический приток, который маскирует гиперполяризационные колебания. Такая возможность кажется вероятной, если учесть, что в некоторых экспериментальных ситуациях ВПСПиТПСП, опосредованные возбуждающими и тормозными вставочными нейронами, могут возникать с примерно равными латентными периодами (Л. Л. Воронин, Л. И. Таненгольц, 1969; Stefanis, Jasper, 1964). Эти наблюдения, следовательно, могут служить указанием на то, что внутреннее торможение складывается где-то в цепи вставочных нейронов условной связи, как это давно предполагал Э. А. Асратян (1970а), и в зависимости от знака условной реакции нейрона формируется различными комбинациями облегчения и подавления активности интернейронов возбуждающего и тормозного действия.
Возможен и другой механизм угасательного торможения без гиперполяризации мембраны. Проведенный Kandel с сотрудниками (1970), Kandel (1970) детальный анализ угашения моносинаптических ВПСП мотонейронов абдоминального ганглия аплизии из числа контролирующих угашение ориентировочного рефлекса втягивания жабер (рефлекс привыкания) дал весьма важные в этом отношении результаты. Оказалось, что снижение амплитуды таких ВПСП связано с ослаблением возбуждающего синаптического действия, основанного, по-видимому, на уменьшении выброса передатчика из пресинаптических терминалей без активного торможения самого мотонейрона.
Предположение о такого рода механизмах угашения клеточных реакций ранее высказали Spenser и соавторы (1966), Segundo и соавторы (1967), исследовавшие синаптические аналоги рефлекса привыкания на спинальных мотонейронах и клетках ретикулярной формации мозгового ствола кошек.
При всех несоизмеримых различиях в организации центральной нервной системы моллюсков и млекопитающих элементарные клеточные механизмы такого рода пластических сдвигов могут оказаться достаточно сходными. Прямые экспериментальные доказательства этого могут быть получены исследованием синаптических механизмов различных форм внутреннего торможения у высших позвоночных на упрощенных моделях условной связи, сохранивших вместе с тем основные качественные особенности условнорефлекторного акта. Если предполагаемое сходство элементарных механизмов угасательного торможения будет доказано, то угашение может предстать как процесс подлинного размыкания - понятие, которым часто пользовался И. П. Павлов.
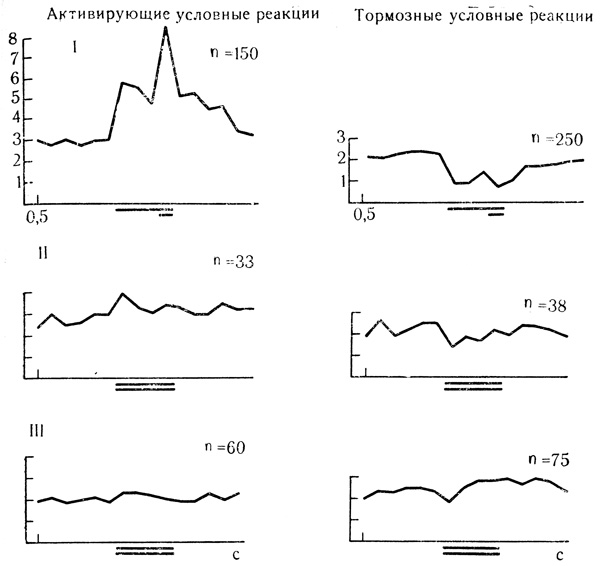
Рис. 58. Динамика выработки дифференцировочного торможения у нейронов двигательной коры с активирующими и тормозными условными реакциями. Активирующие условные реакции: I - усредненные результаты 150 сочетаний для 7 клеток; II - первые 5 проб на дифференцировку для 7 клеток; III - последующие пробы на дифференцировку для 6 клеток. Тормозные условные реакции: I - усредненные результаты 250 сочетании для 8 клеток; II - первые 5 проб на дифференцировку для 8 клеток; III - последующие пробы на дифференцировку для 7 клеток (Е. Л. Полонская, 1974)
Вместе с тем факт активного торможения условных ответов клеток в процессе угашения в виде угнетения разрядов при активирующих реакциях и облегчения - при тормозных постулирует механизмы, которые эту активность поддерживают. Эти механизмы, вероятно, следует искать в остаточной потенциации вставочных клеток возбуждающего и тормозного действия, опосредующих реакции "входных" нейронов с синаптическими входами для сочетаемых стимулов, и, следовательно, их общий гомосинаптический путь на "выходные" нейроны.
Именно потому, что активность вставочных нейронов ослаблена отменой подкрепляющего фактора и по существу является остаточной, угасательное торможение многие авторы описывают как снижение частоты разрядов до исходного уровня фоновой активности. Эффекты торможения и растормаживания разрядов клеток при угашении обычно весьма кратковременны и слабее выражены, чем при дифференцировках, где действуют, по-видимому, иные механизмы поддержания активного характера внутреннего торможения. В пользу такого предположения свидетельствует сравнение усредненных реакций клеток двигательной и слуховой коры на дифференцировочный и угасательный стимулы (рис. 55 и 56, 58 и 59).
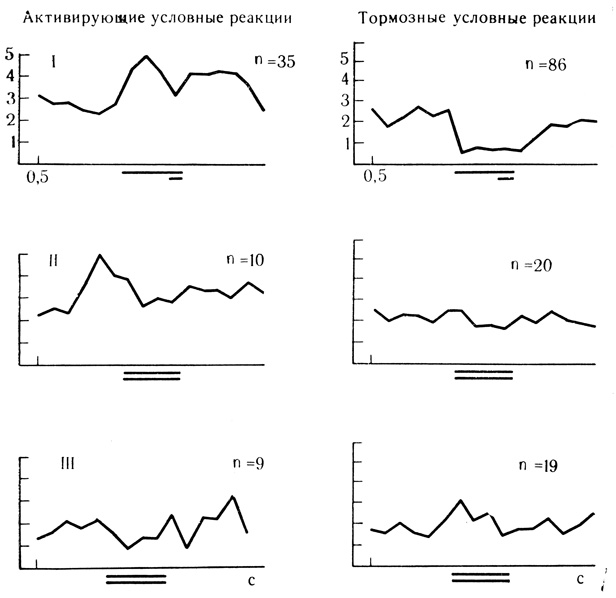
Рис. 59. Выработка дифференцировочного торможения у нейронов слуховой коры с активирующими и тормозными условными реакциями. Активирующие условные реакциии; I - усредненные результаты 35 сочетаний для 2 клеток; II - первые 5 проб на дифференцировку для 2 клеток; III - последующие пробы на дифференцировку для одной клетки. Тормозные условные реакции: I - усредненные результаты 10 сочетаний для 4 клеток; II - первые 5 проб на дифференцировку для 4 клеток; III - последующие пробы на дифференцировку для 3 клеток (Е. Л. Полонская. 1374)
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'