Механизмы воспроизведения следовых процессов условнорефлекторной природы (условные реакции нейронов на время)
Условная реакция нейронов на время оказалась удобной и сравнительно простой моделью для исследования механизмов организации следовых явлений и их воспроизведения на клеточном уровне. Такие реакции вырабатываются при предъявлении сенсорных раздражителей (световых, звуковых, электрокожных) или их сочетаний в фиксированные интервалы времени. За условные реакции на время принимается активность клетки в виде разрядов, опережающих ответы на сенсорную стимуляцию, или разрядов при периодических пропусках раздражителя в заданных временных интервалах.
Опережающие стимул разряды были обнаружены в нейронах гиппокампа при предъявлении световых и звуковых раздражителей с 3-секундным интервалом. Опережающие реакции возникали постепенно, по мере повторения раздражений, определялись интервалами между ними и, следовательно, квалифицированы как реакции на время (О. С. Виноградова, 1965).
В зрительной коре бодрствующих кроликов при повторной стимуляции вспышками света с фиксированными интервалами (около 2 с) реакции на время проявили 5 клеток из 47. Три нейрона ответили разрядом во время пропусков раздражителя в 2-секундном интервале, а 2 клетки реагировали опережающими реакциями ("эффект экстраполяции") (А. Багдонас и др., 1968). Такого рода следовые процессы рассматриваются как нервная модель стимула, которая формируется повторными раздражениями (Е. Н. Соколов, 1969).
Своеобразную реакцию на время нейронов слуховой коры кошки (хлоралозовый наркоз) наблюдал Г. А. Вардапетян (1967). Феномен основан на способности нейронов с on - off-реакциями сдвигать off-компонент ответа на постоянную величину, равную его латентному периоду (600 мс в описанных условиях эксперимента), по мере нарастания длительности звукового сигнала (с 7 до 800 мс). Оказалось, что каждая последующая реакция на стимул большей длительности воспроизводит следовой off-разряд предшествующего ответа, который приходится на время появления этого компонента после выключения предыдущего сигнала. Off-реакция на выключение действующего сигнала возникает с тем же постоянным латентным периодом, как вторая пачка разрядов, следующая за первой пачкой следовой природы. Такие следовые разряды могут воспроизводиться на протяжении до 14 с.
Для изучения следовых явлений на основе которых вырабатываются условнорефлекторные связи, и механизмов воспроизведения этих процессов на уровне нейрона более пригодны модели с применением сочетаний раздражителей, которыми обычно пользуются при выработке двигательных условных рефлексов. В нашей лаборатории условные двигательные рефлексы на время вырабатывались у бодрствующих кроликов сочетаниями звуковых или световых стимулов с электрокожным раздражением конечности (порогового для сгибательного движения), предъявляемых с постоянным 30-секундным интервалом. За клеточные условные реакции на время принималась активность нейрона, несколько (до 1 с) опережающая его ответы на сочетаемые стимулы, или активность, возникающая при следующих друг за другом сериях (5-15) пропусков сочетаний в заданных временных интервалах. Наиболее эффективным оказался прием последовательных пропусков ряда сочетаний, который позволил выявить некоторые существенные свойства воспроизведения следов условного и безусловного компонентов сочетания и выработанной связи между ними. Были исследованы условные реакции на время нейронов двигательной коры (Ф. В. Копытова, М. Я. Рабинович, 1967), дорсального и вентрального отделов гиппокампа (Ф. В. Копытова, Л. К. Куликова, 1970, 1971) и миндалевидного комплекса (Ф. В. Копытова, Ю. С. Медникова, 1972).
В двигательной коре из 175 зарегистрированных нейронов условные реакции на время выработало 89 клеток (51%), в том числе активирующие условные реакции на время (см. рис. 43) проявило 54 клетки (61%), тормозные реакции на время (рис. 44) выработало 35 клеток (39%). Это примерно соответствует относительному числу нейронов моторной коры бодрствующих кроликов, проявивших активирующие и тормозные условные реакции в обычной (классической) экспериментальной ситуации выработки двигательных условных рефлексов (см. главу IV). Несколько более высокий процент клеток, проявивших активирующие условные реакции на время, и немного меньший процент клеток с тормозными следовыми реакциями объясняются отбором преимущественно клеток с активирующими реакциями в начале исследования (Ф. В. Копытова, М. Я. Рабинович, 1967).
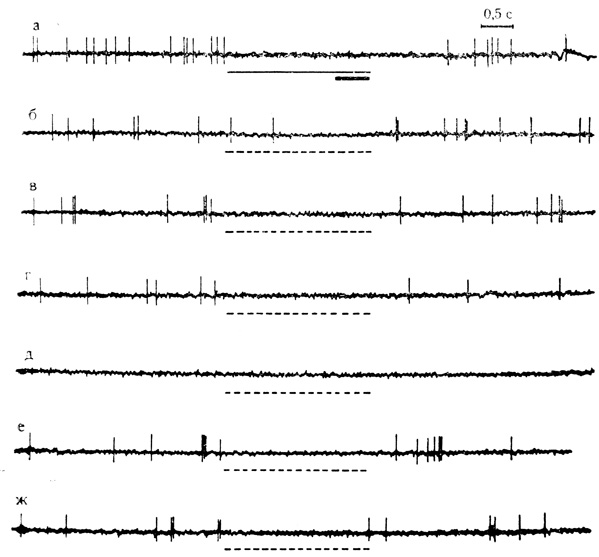
Рис. 44. Тормозные условные реакции на время нейрона двигательной коры. а - 19-е сочетание на клетке, которому предшествовало 146 сочетаний на животном; б - ж - серия из 6 последовательных пропусков сочетаний (пунктирные линии) с 30-секундными интервалами. Следовому торможению активности предшествуют небольшие пачки разрядов (в, е, ж)
В дорсальном отделе гиппокампа условные реакции на время проявило 53% клеток (37 нейронов из 70 зарегистрированных), в том числе 63% клеток выработали активирующие и 37% клеток - тормозные реакции.
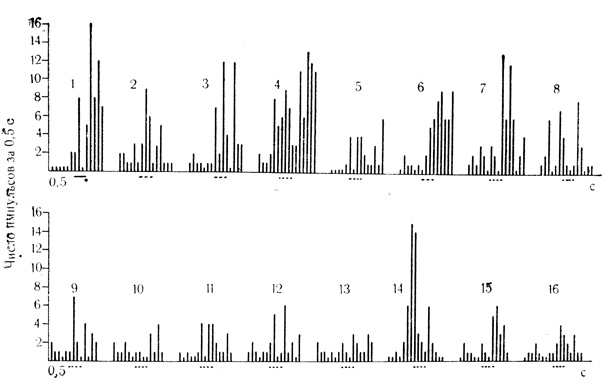
Рис. 45 Активирующие условные реакции на время нейрона дорсального отдела гиппокампа. 1 - 81-е сочетание на клетке, которому предшествовало 300 сочетаний на животном; 2 - 16-я серия из 15 последовательных пропусков сочетаний (пунктирные линии) с 30-секундными интервалами. Наиболее интенсивные следовые разряды возникли во 2-м (3) и 13-м пролусках сочетаний (14); интервалы отставления подкрепляющего стимула воспроизведены во 2-м (3), 8-м (9), 10-м (11) и 11-м пропусках (12)
В вентральном отделе гиппокампа относительное число нейронов, выработавших следовые условные ответы на время, было несколько меньшим - 47% (24 клетки из 51 зарегистрированных) и преобладали активирующие реакции - 58%; тормозные условные реакции на время проявило 42% нейронов. Примеры активирующей и тормозной условных реакций на время нейронов гиппокампа представлены на рис. 45 и 46.
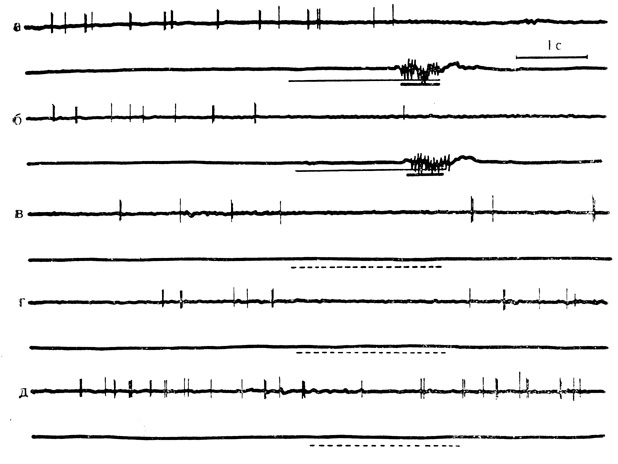
Рис. 46. Тормозные условные реакции на время нейрона дорсального отдела гиппокампа. а и б - 2-е и 19-е сочетания на клетке, которым предшествовало 147 сочетании на животном; в, г, д - серия из 3 последовательных пропусков сочетаний (пунктирные линии) с 30-секундными интервалами после 20-го сочетания. Следовому торможению активности в некоторые пропусках предшествуют небольшие пачки разрядов (Ф. В. Копытова, Л. К. Куликова, 1970)
Преобладание в дорсальном отделе гиппокампа нейронов с тормозными условными реакциями на время следует отнести к структурно-функциональным особенностям поля СА-2 этого отдела, которое расположено наиболее поверхностно, и в котором, по-видимому, была зарегистрирована большая часть клеток, причисленных к дорсальному отделу (Ф. В. Копытова, Л. К. Куликова, 1970). Как показали последующие исследования Ф. В. Копытовой и И. Н. Боголеповой (1972), в поле СА-2 обнаружено 67% клеток с тормозными условными реакциями на время, тогда как в полях СА-3 и СА-4 таких нейронов было только 30%; остальные нейроны этих полей проявили активирующие следовые реакции условнорефлекторной природы (соответственно 33 и 70%). Преимущественно тормозные условные реакции нейронов поля СА-2 можно объяснить преобладанием в этом поле корзинчатых клеток, идентифицированных как вставочные нейроны тормозного действия (Lorente de No, 1934; Eccles, 1969).
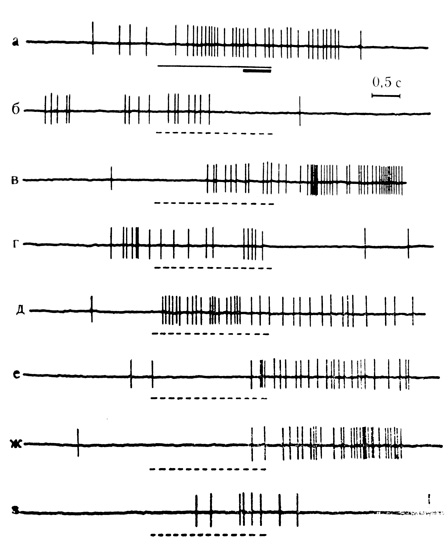
Рис. 47. Активирующие условные реакции на время нейрона миндалины. а - 41-е сочетание; небольшая опережающая реакция клетки на время; б - первый пропуск сочетания; в - з - серия из 6 последовательных пропусков сочетаний, начиная с 5-го. (Во 2-4-м пропусках следовые условные реакции на время отсутствовали.) Наиболее интенсивная следовая реакция возникла в 7-м пропуске (д). Предшествующая разрядам тормозная фаза хорошо выражена в первом пропуске (Ф. В. Копытова, Ю. С. Медникова, 1972)
Из 100 зарегистрированных нейронов миндалевидного комплекса активирующие (рис. 47) и тормозные (рис. 48) условные реакции на время выработало 44 клетки; преобладали нейроны с активирующими следовыми реакциями (60%). В отличие от нейронов двигательной коры и гиппокампа, которые вырабатывали активирующие или тормозные условные реакции в соответствии со знаком действия на клетку подкрепляющего стимула, часть клеток миндалины проявила условные ответы в соответствии с возбуждающим или угнетающим активность действием звукового сигнального раздражителя. По крайней мере из 11 нейронов, которые до сочетаний реагировали на звук, 5 клеток по мере сочетаний выработали стабильные условные реакции на время по знаку действия на клетку сигнального звукового стимула (Ф. В. Копытова, Ю. С. Медникова, 1972).
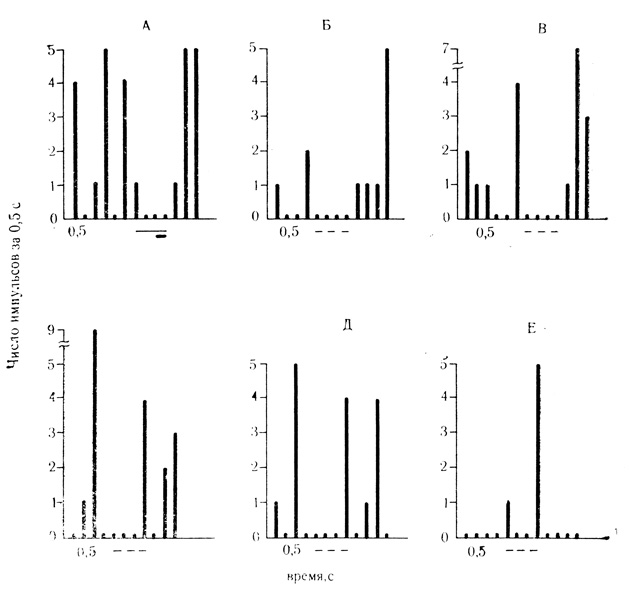
Рис. 48. Тормозные условные реакции на время нейрона миндалины. А - 25-е сочетание; Б-Е - 4, 6, 7, 8 и 9-й пропуски сочетаний. В остальных пропусках (1-3-й и 5-й) условные реакции на время отсутствуют. Тормозным реакциям на время предшествуют отчетливо выраженная активация клетки (Б-Д). В 9-м пропуске (Е) тормозная условная реакция на время угашена; наблюдается растормаживание(Ф. В. Кюпытоза, Ю. С. Медникова, 1972)
Первые условные реакции на время нейронов двигательной коры и гиппокампа в виде опережающих стимуляцию ответов возникали после 10-30 сочетаний одновременно с реакциями на условный сигнал. Для появления стабильных следовых реакций на время в серии последовательных пропусков стимуляции необходимо было примерно 60-70 сочетаний, предъявляемых с фиксированным 30-секундным интервалом. Несколько меньше сочетаний требовали нейроны миндалевидного комплекса: первые реакции на время в виде опережающих ответов появлялись после 4-8 сочетаний, а стабильные следовые реакции в серии последовательных пропусков возникали после 40-50 сочетаний. Другой особенностью нейронов миндалины является постепенное снижение вероятности появления стабильной реакции на время (в среднем на 60-80%) после примерно 100 сочетаний, предъявленных животному по мере выработки условных двигательных рефлексов на время.
Такие свойства нейронов миндалины, как относительно более раннее появление следовых реакций на время и снижение вероятности появления ответов на время по мере упрочения условного рефлекса животного, отражающие динамику условнорефлекторной активности этих клеток, могут быть связаны, как полагают Ф. В. Копытова и Ю. С. Медникова (1972), с функцией миндалевидного комплекса, причастного к выделению мотива-Дионных компонентов сочетания, на что ранее указывали Fuster и Uyeda (1971).
Достаточно стабильные поведенческие реакции животного на время в виде опережающих стимуляцию двигательных рефлексов или движений конечности при пропусках сочетаний появлялись в среднем после 150 сочетаний (рис. 49). При этом каких-либо достоверных корреляций между реакциями нейронов на время и двигательными рефлексами животного не выявлено. В используемой экспериментальной ситуации клеточные следовые реакции во всех исследованных структурах значительно раньше появлялись, были более стабильными и выдерживали больше последовательных пропусков, чем следовые двигательные акты.
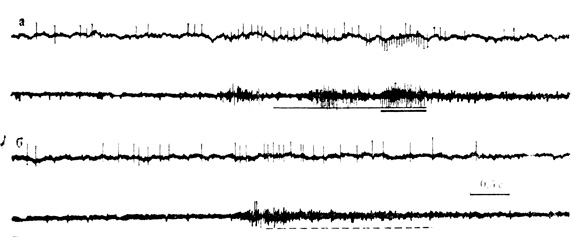
Рис. 49. Двигательные условные реакции кролика на время, а - 140-е сочетание звукового стимула (щелчки 11 в 1 с) с электрокожным раздражением передней конечности. Опережающая мышечная реакция воспроизводит следовой ответ на сигнальный стимул, за которым следует следовая реакция на подкрепление с интервалом его отставления в сочетании и затем - реакция на подкрепляющий стимул; 5 - следовая мышечная реакция ПРИ пропуске раздражителей после 150-го сочетания. На верхней линии каждой осциллограммы - разряды нейрона, на нижней - электромиограмма сгибателей передней конечности (Ф. В. Копытова, М. Я. Рабинович, 1967)
По мере воспроизведения условной активности нейрона на время в серии последовательных пропусков сочетаний постепенно наступает угашение следовой реакции. У нейронов двигательной коры угашение развивается в среднем после 6-8, в гиппокампе - после 15 и в миндалевидном комплексе после 10 пропусков сочетаний. Данные о способности нейронов гиппокампа относительно более длительный период сохранять и воспроизводить следовые реакции согласуются с недавними наблюдениями М. М. Хананашвили (1972): электростимуляция гиппокампа (полей СА-1 и СА-2) приводит к значительному увеличению (в несколько раз) времени, в пределах которого сохраняются временные связи корковых нейронов, выработанные сочетаниями прямого раздражения двух участков поверхности коры. При этом число нейронов, способных вырабатывать такие ассоциации, оставалось неизменным в пределах 20-25%.
Активирующие условные реакции клеток на время; при пропусках раздражений проявлялись либо как первичное воспроизведение следовых процессов в первом пропуске сочетаний (см. рис. 45), либо как отсроченное воспроизведение, когда эффективная реакция возникала в последующих пропусках (см. рис. 43, а также рис. 50), либо имели место обе формы следовой активности (см. рис. 45, 47). Однако отсроченные следовые реакции, как правило, более интенсивны и совершенны по структуре разряда, чем первичное воспроизведение следовых явлений или даже ответы на реальные сочетания. Эти свойства следовых реакций представляются нам весьма существенным аспектом их анализа. Н,а рис. 43 и 50, например, видно, что во 2-м и 3-м пропуске сочетаний соответственно следовые ответы, приуроченные к моменту действия сигнального и подкрепляющего стимулов, воспроизведены значительно более четко, чем это имело место в сочетании, непосредственно предшествовавшем серии пропусков.
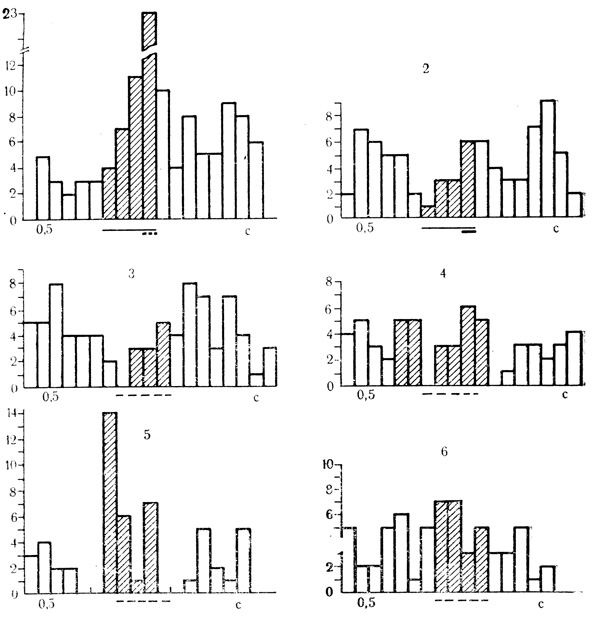
Рис. 50. Отсроченная условная реакция на время нейрона двигательной коры. 1 - пропуск подкрепляющего стимула в 105-м сочетании; 2 - 120-е сочетание звукового стимула с электрокожным раздражением конечности; 3-6 - серия из 4 последовательных пропусков стимула после 120-го сочетания. Интенсивная следовая активация возникает только в 3-м пропуске сочетания (5). Выраженная тормозная фаза, предшествующая следовому разряду нейрона. Реакции нейрона на время и ответы на предъявленные стимулы застрахованы (Ф. В. Копытова, М. Я. Рабинович, 1967)
Нейроны исследованных структур (двигательная кора, гиппокамп, миндалина) с тормозными условными реакциями на время не менее точно воспроизводят следовые ответы при последовательных пропусках сочетаний. При этом, однако, следовое торможение активности в равной степени хорошо выражено во всех пропусках, начиная с первичного воспроизведения в пропуске, следующем непосредственно после сочетания (см. рис. 44, 46, 48).
Отсроченное воспроизведение следовой активности в форме более интенсивных и совершенных реакций рассматривалось нами как клеточный аналог реминисценции - феномена, хорошо известного в психологии памяти (Ф. В. Копытова, М. Я. Рабинович, 1967). В этой связи представляют интерес данные Х. Хидена и П. В. Ланге (1969) о том, что синтез специфических белков в нейронах гиппокампа крыс заметно усиливается к 5-му дню выработки у них двигательных навыков, но достигает оптимального уровня при пробах после 14-дневного перерыва в обучении. В таких отсроченных реакциях, приуроченных к определенному времени, авторы также усмотрели аналогию с явлением реминисценции.
Клеточные аналоги реминисценции можно наблюдать и в других экспериментальных ситуациях, позволяющих воспроизвести следовые реакции на предшествующую стимуляцию. По данным Л. Л. Воронина (19716), полученным на нейронах двигательной коры бодрствующих кроликов с помощью внутриклеточной регистрации, отсроченные (до нескольких десятков секунд) следовые деполяризационные колебания, генерирующие спайковые разряды в ритме предшествовавшего электрокожного раздражения конечности, могут быть более интенсивными, чем первые следовые реакции после прекращения"* стимуляции и ответы на реальный раздражитель (рис. 51, А, Б, В). По длительности, временному течению и реакциям на поляризующий ток (рис. 51, Г) следовые деполяризационные колебания не отличаются от постсинаптических потенциалов, вызванных электрокожным раздражением конечности, и отражают процессы, протекающие главным образом в пресинаптических элементах.
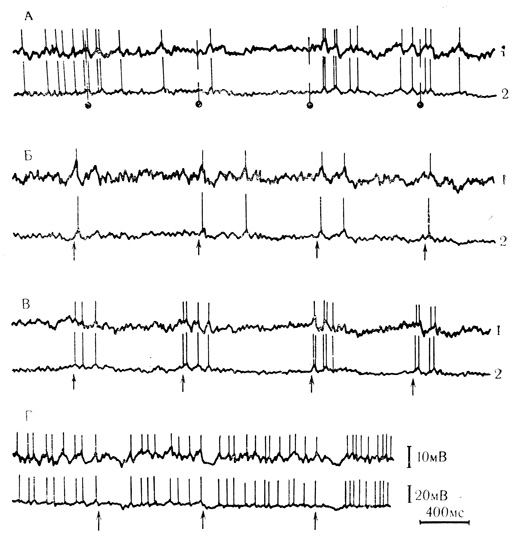
Рис. 51. Следовые деполяризационные колебания, генерирующие спайковые разряды нейрона двигательной коры кролика после ритмического электрокожного раздражения передней конечности. А - постсинаптические реакции в ответ на электрокожное раздражение конечности (отмечены точками); Б и В - следовые постсинаптические реакции (отмечены стрелками) через 20 с (Б) и 48 с (В) после прекращения электрокожного раздражения; Г - следовые реакции того же нейрона при деполяризации током 10-9А через внутриклеточный микроэлектрод. Искусственная деполяризация вызывает увеличение гиперполяризационных компонентов следового ответа, что идентифицирует следовые деполяризационные колебания как постсинаптические реакции. 1 - усилитель переменного тока (RC = 0,1 с); 2 - усилитель постоянного тока (Л. Л. Воронин, 19716)
Сходные результаты на другой экспериментальной модели были получены В. Г. Скребицким и А. Н. Чепковой (1973). Исследовались реакции нейронов двигательной коры бодрствующих кроликов на совпадающие сочетания (с нулевым интервалом) околопороговой стимуляции вентрального задне-латерального ядра таламуса (VPL) и локальной поляризации клеток корковой моторной зоны выходящим током через регистрирующий микроэлектрод. Сочетания вызывали значительное усиление начального разряда и заполнение импульсами последующей тормозной паузы ответов клетки на стимуляцию таламического ядра (рис. 52, А). После выключения поляризующего тока следовая активирующая реакция на одну таламическую стимуляцию появлялась не сразу, а спустя 30-50 с, на 2-3-е предъявление (рис. 52, Б).
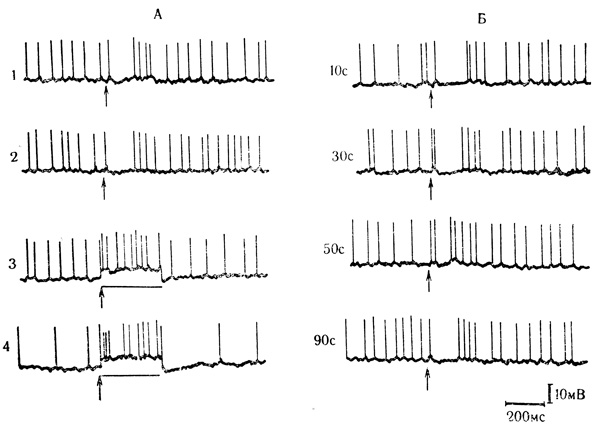
Рис. 52. Следовые постсинаптические реакции нейрона двигательной коры в последействии совпадающих сочетаний стимуляции вентрального задне-латерального ядра таламуса (VPL) с локальной активирующий поляризацией клетки через микроэлектрод. А: 1, 2 - реакции нейрона на околопороговую стимуляцию VPL в виде начального разряда с последующей тормозной паузой и посттормозной отдачей; 3, 4 - сочетания стимуляции VPL (стрелки) и поляризации (300 мс, отмечена горизонтальной линией); Б - после выключения поляризующего тока клетка реагирует на стимуляцию VPL следовым усилением начального разряда при 2-м предъявлении стимула (через 30 с) и заполнением импульсами тормозной паузы при 3-м предъявлении стимула - через 50 с (В. Г. Скребицкий, А. Н. Чепкова, 1973)
Обращает на себя внимание фаза торможения активности нейронов длительностью до 1 с, которая часто предшествует следовым разрядам клетки на время. Эту предварительную тормозную фазу можно видеть почти на всех осциллограммах (см. рис. 43 и 47), но особенно отчетливо она бывает выражена на гистограммах, например, на рис. 50, 5. При анализе 210 следовых разрядов клетки у 27 нейронов двигательной коры предварительная фаза торможения была отмечена в 51% случаев, в том числе: в 31% случаев частота разрядов нейрона уменьшилась по сравнению с фоновой активностью в 2 раза, в 14% случаев - в 3 раза, в 6% случаев - в 6 раз. В 13% случаев предшествующее уменьшение частоты разрядов было столь незначительным, что принималось за случайные колебания активности. В 36% случаев предшествующего снижения частоты разрядов клетки не было (Ф. В. Копытова, М. Я. Рабинович, 1967).
Складывается впечатление, что следовая активация на время возникает как посттормозная отдача или па крайней мере усиливается предварительным торможением. Наряду с этим тормозным следовым реакциям на время также часто предшествует некоторое повышение активности в виде небольшой пачки импульсов (см. рис. 44 и 46), которое особенно отчетливо выражено на гистограммах рис. 48.
Нам представляется, что предшествующие следовым эффектам короткая фаза торможения при активирующих реакциях и фаза активации при тормозных следовых ответах заслуживают специального экспериментального анализа с использованием точных количественных Следует подчеркнуть, что все нейроны исследованных структур (двигательная кора, гиппокамп, миндалина), выработавшие условные связи при сочетаниях звуковых стимулов с электрокожным раздражением конечности, были способны проявлять условные реакции на время в виде следовой активности в серии последовательных пропусков сочетаемых раздражителей.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'