"Сцепление" следов условного и безусловного стимулов
При выработке условных связей на уровне нейрона механизмы "сцепления" следов условного и безусловного стимулов проявляют себя при пропусках в отдельных сочетаниях подкрепляющего раздражителя и при пропусках сочетания в целом в условных реакциях клеток на время. Известно, что при однократном пропуске подкрепления в сложившейся условной связи сигнальный стимул становится способным не только вызвать ответ нейрона, но и воспроизвести его следовую реакцию на подкрепление с интервалом предшествующего отставления в сочетаниях (Л. Л. Воронин, Р. Г. Кожедуб, 19716; Е. Л. Полонская, М. Я. Рабинович, 1971; Adam e. a., 1971; Adorjani e. a., 1971). Этот феномен настолько адекватно отражает выработанную связь между сочетаемыми стимулами, что используется как прием верификации подлинности условных клеточных реакций (Л. Л. Воронин, Р. Г. Кожедуб, 1970, 1971; Е. Л. Полонская, М. Я. Рабинович, 1972; Rabinovich, Polonskaya" 1973; Adorjani e. a., 1971).
На синаптическом уровне воспроизведение связанных с условной реакцией следов подкрепления было показано Л. Л. Ворониным и Р. Г. Кожедуб (19716). Деполяризационные колебания, генерирующие спайковые разряды в ответ на подкрепление, сохраняются и при его пропуске в сочетаниях. При этом частота следовых ВПСП может соответствовать частоте импульсов безусловного раздражения больше, чем это обычно имеет место при действии самого стимула. Примеры воспроизведения следовых реакций на подкрепляющий стимул представлены на рис. 42.
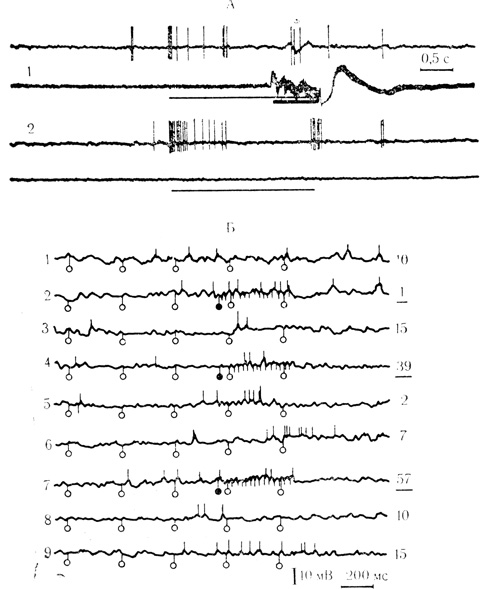
Рис. 42. Следовые реакции нейронов на подкрепляющий стимул при его пропусках в сочетании. А - внутреннее коленчатое тело. 1 - 18-е сочетание звукового стимула с электрокожным раздражением конечности; 2 - последующий пропуск подкрепления, активирующая следовая реакция (Е. Л. Полонская, 1Э74). Б - двигательная кора. Клеточный аналог условного рефлекса вырабатывался сочетаниями электрического раздражения двух пунктов корковой поверхности. Квазивнутрнклеточная запись. 1 - возбуждающая синаптическая реакция клетки на 'условный' стимул (светлые кружки) до сочетаний. Показаны реакции на последние 5 импульсов из 7 составляющих 'условный' стимул; 2, 4, 7 - 1-я, 2-я и 3-я серии сочетаний соответственно; 3, 5, 6, 8, 9 - угашение условной реакции в каждой из серий. Подчеркнутые цифры справа от осциллограмм - порядковый номер сочетания, не подчеркнутые - пробы на угашение. Следовая синаптическая реакция на пропущенное подкрепление в некоторых пробах (сравните 7 и 9) интенсивнее ответа на реальный стимул (Л. Л. Воронин, Р. Г. Кожедуб, 19716)
Механизмы воспроизведения следов сигнальной и подкрепляющей стимуляции при выработке условных реакций клеток на время рассматриваются в следующем разделе главы. Здесь, однако, уместно отметить, что при последовательных пропусках ряда сочетаний в фиксированных интервалах их предъявлений следовые реакции, соответствующие предшествовавшим условному и безусловному стимулам, часто возникают с интервалами между ними в пределах отставления подкрепляющего раздражителя (рис. 43, г; см. также рис. 45, 3,5).
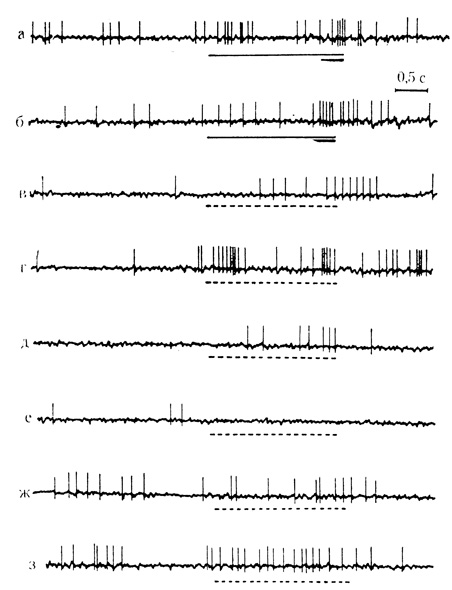
Рис. 43. Активирующие условные реакции на время нейрона двигательной коры. а и б - 45-е и 50-е сочетание звукового стимула (щелчки 15 в 1 с) с злектрокожным раздражением конечности; условный стимул обозначен тонкой линией, подкрепляющий - толстой; в - з - серия из 6 последовательных пропусков сочетаний (пунктирная линия) с 30-секундными интервалами. Наиболее интенсивные следовые разряды с воспроизведением интервала отставления подкрепляющего стимула в сочетании, возникли во втором пропуске (г) после 50-го сочетания
Подтверждением реальности механизма "сцепления" следов стимуляции могут служить также результаты исследования эффектов сочетания на фоне поляризации двух раздражителей различной сенсорной модальности, проведенные Morrell (1967). При анодной поляризации поверхности коры нейроны зрительной зоны реагируют на световые и звуковые (или световые и электрокожные) стимулы с различной и постоянной латентностью. В процессе одновременного предъявления этих стимулов появляются ответы, в которых либо сохраняются два максимальных разряда, соответствующих латентным периодам сочетаемых раздражителей, либо возникает новая структура разряда, с иным распределением импульсации во времени. После 80-100 таких сочетаний нейрон приобретает способность воспроизводить некоторое время всю структуру предшествующего ответа действием одного светового стимула.
В классических условных рефлексах механизмы "сцепления" следов раздражений проявляют себя постоянно и в самых разнообразных формах, что позволяет ограничиться немногими примерами. Наиболее ярко феномен "сцепления" следов выступает в динамической стереотипии. Такая система складывается повторными предъявлениями стереотипа положительных и отрицательных раздражителей одной или различной модальности и после ее упрочения может быть целиком воспроизведена только первым из стимулов, сохраняя при этом качественные и количественные особенности реакции на каждый из компонентов стереотипа раздражений (Э. А. Асратян, 1938). ЭЭГ-исследования динамической стереотипии показали, что сложившиеся особенности электрографических реакций структур мозга на каждый из стимулов стереотипа точно воспроизводятся как чисто следовое явление при экстренной отмене всего комплекса реальных раздражителей (П. К. Анохин, 1957).
В 30-х годах П. С. Купаловым был обнаружен весьма примечательный факт, что сокращение длительности действия сигнального стимула (например, с 30 до 3-5 с) не сказывается на проявлении условного слюноотделения при реализации наиболее простого условного рефлекса с пищевым подкреплением. Более того, при последующих специальных исследованиях оказалось, что первоначальное 30-секундное действие условного звукового стимула может быть сокращено до 0,1 с и даже до 0,01 с без заметного уменьшения секреторной реакции на протяжении всех 30 с, предшествующих пищевому подкреплению (П. С. Купалов, 1947а).
Основываясь на этих фактах, можно полагать, что для последовательного развертывания отдельных компонентов и более сложных поведенческих актов также достаточен только начальный разряд в афферентных путях, который способен запустить всю систему "сцепленных" следов (энграмму поведенческого акта) предшествующих раздражений. Такой механизм отчетливо демонстрируют ситуационные условные рефлексы П. С. Купалова, которые включают большой ряд разнообразных сенсорных стимулов и ответных двигательных актов, расположенных в определенной временной и пространственной последовательности и завершающихся пищевым подкреплением. Когда ситуационные рефлексы достаточно упрочены, вся последовательность точно скоординированных двигательных актов воспроизводится действием первого из сенсорных стимулов (П. С. Купалов, 1963; П. С. Купалов и др., 1964).
О том же свидетельствуют механизмы становления и реализации цепных пищедобывательных условных рефлексов, которые состоят из ряда пищедобывательных двигательных актов, выработанных на различные сенсорные стимулы в определенной последовательности их включения. После упрочения цепных условных рефлексов весь ряд двигательных актов этой сложной реакции легко воспроизводится действием первого в ряду сенсорного стимула (Л. Г. Воронин, 1967).
Механизмы "сцепления" следов лежат, вероятно, в основе и так называемых ассоциативных условных связей: после предварительных сочетаний двух индифферентных сенсорных стимулов и выработки в последующем на один из них пищевого или оборонительного условного рефлекса второй раздражитель в равной степени приобретает сигнальное значение для соответствующего рефлекса. Интересно, что способность вырабатывать такого рода связи появляется в ряду позвоночных, начиная с рептилий (Б. Ф. Сергеев, 1967; А. И. Карамян, 1970). Другой пример вырабатываемой ассоциации между двумя раздражителями приведен в исследовании Э. А. Костандова и Ю. Л. Арзуманова (1973), проведенном на людях. При последовательном предъявлении двух зрительных стимулов (стрелка и слово на светящемся табло) с односекундным интервалом между ними в затылочной области регистрируются суммарные ответы на каждый из них. После ряда таких сочетаний между стимулами образуется связь, которая проявляет себя при пропусках второго стимула. Следовая реакция на второй стимул возникает в пределах того же односекундного интервала, как это обычно имеет место при пропусках в сочетании подкрепления. Однако и здесь ассоциация могла быть выработана, если второй стимул (слово на табло) вызывал у испытуемого эмоциональную реакцию (например, напоминание о неприятной в прошлом ситуации).
Нам представляется, что только механизмами "сцепления" следов предшествующей условной и безусловной стимуляции могут быть удовлетворительно объяснены хорошо известные факты воспроизведения действием электрического тока сложных двигательных актов, в том числе инструментальных, выработанных сочетаниями электрораздражений сенсорных и моторных структур мозга. Реализация таких условнорефлекторных актов немыслима без предварительной выработки их энграмм, т. е. определенным образом организованных в пространстве и времени следовых процессов (Н. Ю. Беленков, 1973; Doty, 1969).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'