Сочетание локальной поляризации отдельных нейронов двигательной коры с действием звуковых стимулов
У 11 бодрствующих кроликов зарегистрирована импульсная активность ПО нейронов двигательной коры в зоне проекции передней конечности. При этом были исследованы реакции клеток на локальную поляризацию анодным током, выходящим через внеклеточный регистрирующий микроэлектрод, и на сочетания активирующей поляризации нейрона с действием звуковых стимулов. Вероятные механизмы возбуждающего действия на нейроны выходящего из микроэлектрода поляризующего тока обсуждались в ряде работ. По-видимому, входящий в клетку ток локально гиперполяризует участок мембраны под микроэлектродом, тогда как другие участки, включая низкопороговую область начального сегмента аксона, деполяризуются выходящим из клетки током (Л. Л. Воронин, 1966, 1968а; Strumwasser, Rosenthal, 1960; Gerin, Courjon, 1962; Bindman e. a., 1964; Spehlmann, Kapp, 1964, и др.).
Оптимальные значения поляризующего тока подбирали для каждого нейрона по степени его активирующей реакции и колебались в пределах от 0,1 до 2*10-9 А. Цепь поляризации замыкалась через серебряную пластинку, укрепленную на ухе животного. Поляризация продолжалась от 3 до 30 мин. В качестве звуковых стимулов применяли тоны с частотой 200 и 500 кол/с длительностью 0,5 с. Тон 200 кол/с ("положительный" стимул) многократно (до 35 раз) предъявлялся на фоне поляризации клеток; тон 500 кол/с ("отрицательный" стимул) применялся во время поляризации только 1-2 раза (Ф. В. Копытова, М. Я. Рабинович, 1970).
Из 110 нейронов на действие локальной поляризации йодным током реагировало 100 клеток (91%), в том числе, как это следует из табл. 10, у 85 клеток (77%) локальная поляризация вызвала повышение частоты разрядов, у 15 нейронов (около 14%) - торможение активности; 10 клеток (около 9%) на включение постоянного тока не реагировали.
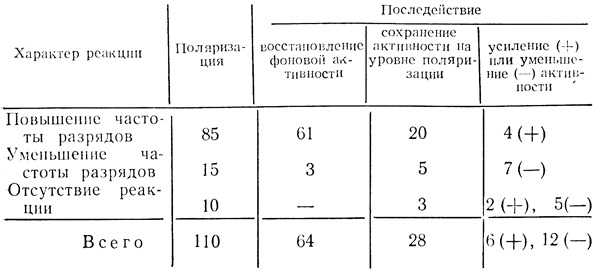
Таблица 10. Реакции нейронов двигательной коры на локальную поляризацию анодным током и в последействии
После выключения поляризующего тока у 64 нейронов (58%) восстановилась исходная частота фоновых разрядов, 25 клеток сохранили в последействии эффекты поляризации, 6 клеток проявили в последействии повышение частоты разрядов, а 12 клеток - уменьшение им-пульсации по сравнению с исходной фоновой активностью. Следовые эффекты поляризации, таким образом, проявили только 43 нейрона (39%). Выключение поляризующего тока сказалось также на активности тех нейронов, которые на поляризацию не реагировали. Из 10 таких клеток у двух в последействии разряды усилились, а у 5 нейронов отчетливо снизились. Чтобы исследовать способность клеток дифференцировать звуковые стимулы в зависимости от повторности их предъявления на фоне локальной поляризации, отбирали преимущественно нейроны, которые до поляризации на эти стимулы не отвечали. Зарегистрирована 71 такая клетка.
Были также использованы результаты анализа активности 39 нейронов, которые в контрольных пробах реагировали на оба (30 клеток) или один (9 клеток) из предъявляемых звуковых стимулов.
Включение поляризующего тока изменило функциональное состояние почти всех клеток, не реагировавших в контрольных пробах на звук. На фоне поляризации около 82% нейронов стало отвечать на оба стимула, около 15% - на один стимул. Значительная часть клеток сохранила способность к реакциям на звук и после выключения поляризующего тока (табл. 11). К нейронам, отвечающим на звук при поляризации и в последействии, мы относили клетки, которые отвечали на стимуляцию не менее чем в 50% проб.

Таблица 11. Распределение нейронов двигательной коры по их ответам на звуковые стимулы 'положительного' и 'отрицательного' значения при локальной поляризации и в последействии
Примечание. В скобках указано количество клеток, которые на звуковые стимулы реагировали торможением активности.
Из табл. 11 следует, что 22 нейрона из 58 клеток, которые на фоне поляризации начали реагировать на оба звуковых стимула ("положительный" и "отрицательный"), после выключения поляризующего тока сохранили ответы только на стимул положительного значения. Для более полного учета клеток, способных в последействии поляризации различать раздражители в зависимости от предшествующего их применения на фоне поляризации, к 58 нейронам необходимо прибавить 14 клеток (из 30 отвечавших на звук в контрольных пробах), у которых при поляризации заметно усилились реакции на оба стимула. До поляризации реакции этих клеток на звук были нестойкими и некоторое повышение частоты разрядов в ответ на стимул имело место не более чем в 40% проб. На фоне поляризации эти клетки реагировали на звук в 80-90% проб с частотой импульсов, почти в 2 раза превышавшей частоту разрядов в контрольных пробах. Из 14 таких нейронов 2 клетки сохранили способность в последействии отвечать только на "положительный" звуковой стимул.
Таким образом, из 72 нейронов, которые на фоне поляризации отчетливо реагировали на оба стимула, 24 клетки в последействии поляризации отвечали только на "положительный" стимул. Следовательно, около 33% клеток проявили способность дифференцировать "положительный" звуковой стимул, многократно применявшийся на фоне их поляризации через внеклеточный микроэлектрод, от "отрицательного" стимула, который предъявлялся в аналогичных условиях не более 1-2 раз.
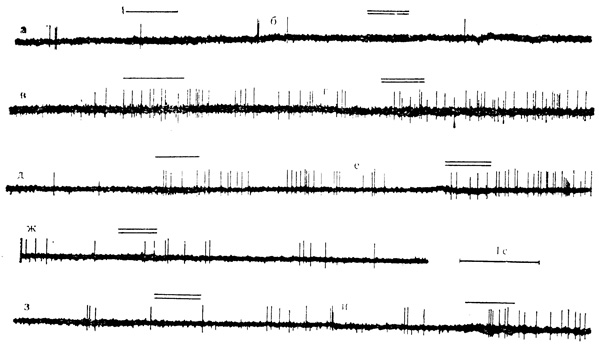
Рис. 39. Реакции нейрона двигательной коры на 'положительный' и 'отрицательный' звуковые стимулы при их сочетании с локальной поляризацией через микроэлектрод и в последействии. Клетка на глубине 1350 мкм. а и б - контрольное предъявление 'положительного' и 'отонцательного', звуковых стимулов; в и г - на фоне поляризации (0,4*10 А) нейрон отвечает на оба стимула (16-я и 18-я минута поляризации); д - и - в последействии поляризации клетка реагирует вначале на оба стимула (д, е - 1-я минута последействия), затем ответ на 'отрицательный' стимул заметно уменьшается и исчезает (ж и з - 2-я минута последействия). Ответ на 'положительный' стимул (и - 6-я минута последействия) сохранен (Ф. В. Копытова, М. Я. Рабинович, 1970)
На рис. 39 и 40, А, Б, В видно, что нейроны, которые в контрольных пробах не реагировали на оба звуковых стимула, начинают отвечать на эти раздражители во время действия поляризующего тока повышением частоты разрядов. После выключения поляризации эти клетки на протяжении нескольких минут реагируют только на "положительный" стимул и не отвечают на звуковой раздражитель "отрицательного" значения. отдельных случаях способность дифференцировать "положительные" и "отрицательные" стимулы в последействии поляризации возникает через некоторое время (1-3 мин) после выключения поляризующего тока. Такая отсроченная дифференцировка "положительного" стимула представлена на рис. 39 и 40, Б.
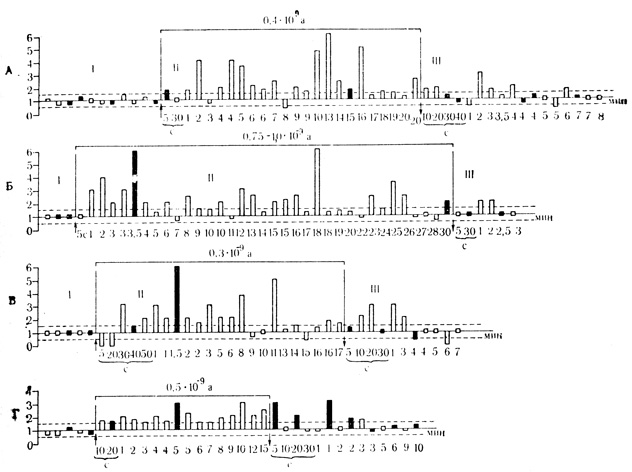
Рис. 40. Динамика ответов клеток двигательной коры на 'положительный' и 'отрицательный' звуковые стимулы на фоне локальной поляризации через микроэлектрод до и после выключения поляризующего тока. А - спонтанно активный нейрон на глубине 880 мкм; Б - спонтанно неактивный нейрон на глубине 1620 мкм. Поляризующий ток 1,0-10 А включен 10-й минуте после начала поляризации током 0,75-10 А; В - спонтанно реактивный нейрон на глубине 1096 мкм; Г - спонтанно активный нейрон на глубине 1020 мкм. В последействии поляризации клетка проявила парадоксальную реакцию. Дополнительные пояснения см. на рис. 38 (Ф. В. Копытова, М. Я. Рабинович, 1970)
Обращает на себя внимание то, что некоторые нейроны способны различать "положительный" и "отрицательный" звуковые стимулы и тогда, когда эти раздражители вызывали на фоне поляризации торможение спонтанной активности. Такие клетки (2 нейрона) после выключения поляризующего тока отвечали на "положительный" стимул торможением импульсации. "Отрицательный" стимул подобной реакции не вызвал (рис. 41).
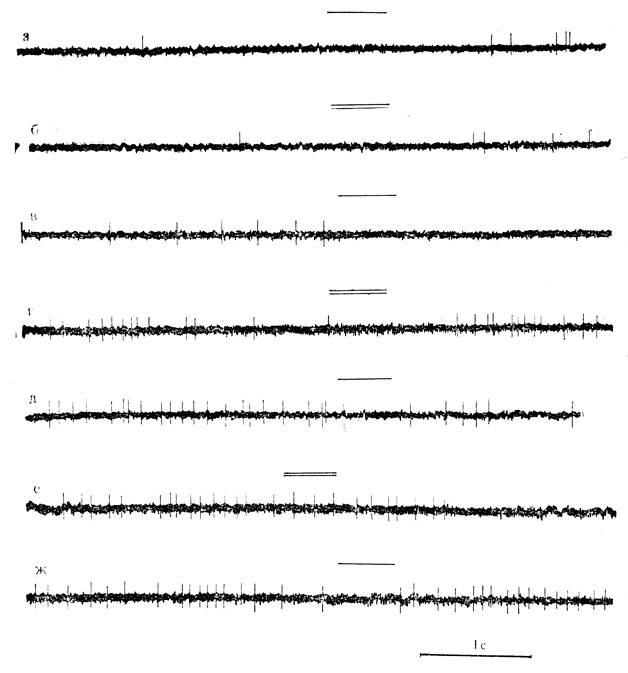
Рис. 41. Воспроизведение тормозной реакции клетки двигательной-коры на 'положительный' звуковой стимул в последействии поляризации. Нейрон на глубине 1420 мкм. а и б - контрольное предъявление 'положительного, и 'отрицательного' стимулов; в и г - на фоне поляризации (0,5*10-9 А) оба стимула вызывают торможение разрядов (6-я минута поляризации); д - ж - после выключения поляризующего тока клетка реагирует торможением на 'положительный, стимул (д - 1-я минута ж - 4-я минута последействия) и слабо реагирует на 'отрицательный' - стимул (е - 1-я минута последействия) (Ф. В. Копытова, М. Я. Рабинович 1970)
Шесть нейронов проявили парадоксальную реакцию и в последействии поляризации некоторое время отвечали только на стимул отрицательного, значения, который с поляризацией повторно не сочетался (рис. 40, Г).
Дифференцированные ответы на "положительный" и "отрицательный" стимулы могли сохраняться в последействии локальной поляризации нейронов через микро-электрод максимально до 14 мин.
Из 24 клеток, проявивших способность дифференцировать "положительный" и "отрицательный" звуковые-раздражители, 22 клетки сохранили это свойство до 1 мин, 15 клеток - до 5 мин, 5 клеток - до 10 мин и 2 клетки - до 14 мин. Парадоксальные реакции сохранялись не более 3 мин.
Как и при суммарной поляризации, величина ответов клеток на оба звуковых стимула при их локальной поляризации заметно варьировала от пробы к пробе. Однако и здесь большинство ответов превышало уровень, принятый нами за значимый (см. рис. 40).
При сравнений эффектов локальной поляризации отдельных нейронов двигательной коры с суммарной поляризацией популяций клеток той же корковой зоны, обращает на себя внимание ряд существенных различий, между ними. На локальную поляризацию через регистрирующий микроэлектрод реагировал 91% нейронов, в; том числе 77% клеток проявили повышение частоты разрядов, а 14% - торможение активности. Суммарная поляризация через макроэлектрод, расположенный на поверхности коры, вызвала реакцию у 75% клеток, в том числе 60% отвечали активацией, а 15% - торможением разрядов. На фоне локальной поляризации 82%. клеток приобрели способность реагировать на ранее не* деятельные звуковые стимулы (либо ответы усиливались и стабилизировались), при суммарной поляризации на звуковые стимулы стало отвечать только 67% клеток.
Вместе с тем при суммарной поляризации следовые эффекты поляризующего тока сохранялись некоторое время в последействии либо даже усиливались у 49% клеток, тогда как при локальной поляризации такого рода пластические сдвиги имели место у 39% клеток. При суммарной поляризации 47% нейронов сохранили в последействии способность различать "положительные" и "отрицательные" звуковые стимулы на основе "сцепления" следов предшествующих сочетаний поляризующего тока с действием "положительного" звукового раздражителя, при локальной поляризации таких клеток было только 33%.
Эти факты указывают на то, что для появления у нейронов двигательной коры таких функциональных свойств, как способность реагировать на ранее индифферентные сенсорные стимулы, достаточна деполяризация мембраны клеточных тел, которая более эффективна при локальной поляризации нейрона через микроэлектрод. Полагают, что деполяризация облегчает реакцию клетки на подпороговые ВПСП, вызванные сенсорной стимуляцией (Л. Л. Воронин, 1966, 1968а; Spehlmann, 1963; Spehlmann, Карр, 1964).
Для образования относительно устойчивой связи на уровне нейрона непосредственная деполяризация клеточных тел явно недостаточна. По данным Л. Л. Воронина и Е. И. Солнцевой (1969а, б), следовые эффекты поляризующего тока, как правило, не связаны с изменениями уровня деполяризации или гиперполяризации мембраны нейрона. По-видимому, для реализации механизма "сцепления" следов необходимо воздействие дополнительных факторов. К такого рода факторам следует отнести вторичные синаптические влияния со стороны других клеток во взаимодействующей системе нейронов, одновременно подвергнувшихся поляризации при воздействии анодного тока на поверхность коры (Л. Л. Воронин, 1968а; Purpura, McMurtry, 1965; Gorman, 1967; Lippold, 1970), затем значительное повышение эффективности синаптического влияния на такие клетки сенсорной стимуляции (звук, свет), которое связывается с активацией дополнительных (резервных) вставочных нейронов, и, наконец, повышение возбудимости дендритов, которое предположительно имеет место при суммарной поляризации популяции нейронов (Л. Л. Воронин, 1968а).
Эффекты суммарной поляризации реализуются, таким образом, механизмами, которые следует иметь в виду при нейрофизиологическом анализе взаимосвязи условного и безусловного стимулов. По-видимому, эти механизмы в какой-то степени имитируют действие на клетку подкрепляющего рефлекса и включаются сочетаниями при выработке условных реакций на уровне нейрона. В пользу такого предположения свидетельствуют почти равные количества нейронов двигательной коры, проявивших способность различать "положительные" и "отрицательные" стимулы при суммарной поляризации клеток, и нейронов, способных вырабатывать условные связи сочетаниями натуральных раздражителей (соответственно 47 и 52%).
В заключение необходимо отметить, что как при активирующей суммарной поляризации, так и при локальном воздействии активирующего тока в последействии имели место тормозные следовые эффекты. Последние могут быть объяснены двойным действием поляризующего тока любого направления, деполяризующего одни участки клеточной мембраны и гиперполяризующего другие, и степенью выраженности адаптивных свойств нейрона к поляризующему току, которые определяют знак следовой реадаптации (Л. Л. Воронин, Е. И. Солнцева, 1969а, б). Не менее важным фактором, определяющим знак следовой реакции, являются, по-видимому, вторичные воздействия поляризующего тока на синаптический аппарат клетки с преимущественным использованием либо возбуждающих, либо тормозных путей (Lippold, 1970). Этим обстоятельством можно объяснить весьма важный факт "сцепления" следов не только (активирующего, но и тормозного значения.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'