Эффекты сочетания суммарной поляризации популяции нейронов двигательной коры с действием звуковых стимулов
Зарегистрирована активность 108 нейронов двигательной коры (14 бодрствующих кроликов) до поляризации анодным током поверхности этой области, на фоне поляризации и после выключения поляризующего тока Поляризующий электрод (толстая нить, смоченная в физиологическом растворе, или серебряная проволока диаметром 0,3 мм) помещали на поверхности коры рядом с регистрирующим микроэлектродом. Цепь поляризации замыкалась через металлический зажим на ухе животного. После обнаружения клетки определяли ее реакции на звуковые стимулы, а затем на 10-35 мин включали анодный ток, величину которого подбирали до получения отчетливой реакции клетки на звук (1-10 мкА). Активность нейронов и их ответы на стимулы регистрировали на протяжении всего времени поляризации и до 20-30 мин после выключения поляризующего тока. Повторную поляризацию производили в большинстве случаев через 40 мин. В качестве звуковых стимулов использовали тоны с частотой 150-1500 кол/с.
В каждом опыте применяли два различных по частоте тона, один из которых ("положительный") многократно (до 20-25 предъявлений) использовался на фоне поляризации, а другой ("отрицательный") предъявлялся 1-2, а иногда и 3 раза (М. Я. Рабинович, Ф. В. Копытова, 1969).
Из 108 зарегистрированных нейронов на действие поляризующего анодного тока реагировало 82 клетки (75%), в том числе у 65 клеток (около 60%) анодная поляризация вызывала повышение частоты разрядов, у 17 клеток (около 15%) - торможение активности; 26 нейронов (около 24%) на поляризацию поверхности коры (в пределах примененных величин тока и длительности его действия) не реагировало (см. табл. 8). Реакции возникали с вариабельной латентностью в пределах от 40 мс до нескольких сотен миллисекунд.
Как следует из табл. 8, после выключения поляризующего тока только у 46 нейронов (44%) восстановилась исходная фоновая активность. У 53 нейронов (49%) на протяжении 20-30 мин в последействии сохранялись (24 клетки) либо даже усиливались активирующие (15 клеток) и тормозные (14 клеток) следовые эффекты поляризации.

Таблица 8. Реакции нейронов двигательной коры на суммарное действие поляризующего тока и в последействии
В том числе 18 нейронов относились к группе из 26 клеток, которые на поляризующий ток не реагировали: 12 клеток проявили в последействии поляризации следовое повышение частоты разрядов, а 6 клеток - торможение активности.
Для анализа эффектов сочетания поляризации поверхности коры с действием звуковых стимулов различной частоты отбирали преимущественно нейроны, не реагировавшие на звуковые стимулы. Из 179 опробованных по этому поводу нейронов была отобрана 91 клетка. Для сравнения результатов сочетаний было исследовано также 17 нейронов, которые до Поляризации реагировали на один или оба звуковых стимула. Из 91 нейрона двигательной коры на фоне анодной поляризации ее поверхности стало отвечать на оба звуковых стимула - "положительный" и "отрицательный" - 45 клеток; только на "положительный" стимул реагировало 13 клеток; только на "отрицательный" стимул - 2 клетки; 31 нейрон не отвечал на область раздражителя. Следовательно, около 67% клеток начали реагировать повышением частоты разрядов на ранее недругтельные звуковые раздражители.
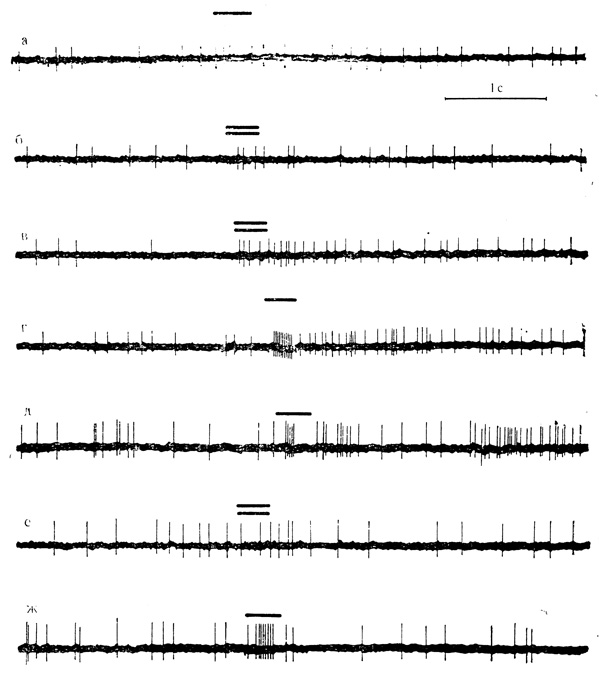
Рис. 37. Реакции нейрона двигательной коры на 'положительный' и 'отрицательный' звуковые стимулы при их сочетании с суммарной поляризацией корковой поверхности и в последействии. Клетка на глубине 1470 мкм. До поляризации клетка на 'положительный' (а - тон 300 кол/с, одна черта) и 'отрицательный' (б - тон 1500 кол/с, две черты) стимулы почти не реагирует. На фоне поляризации (1,5 мкА) клетка отвечает на оба стимула хорошо выраженными разрядами (в - 7-я минута действия поляризующего тока; г - -я минута поляризации). В' последействии поляризации сохраняются ответы на 'положительный' стимул (д - 1-я минута последействия; ж - 5-я минута) и отсутствуют на 'отрицательный' (е - 1-я минута) стимул (М. Я. Рабинович, Ф. В. Копытова, 1969)
Примеры таких клеток представлены на рис. 37, а-г и на рис. 38. К нейронам, реагирующим на звуковое раздражение, мы относили только те клетки, которые отвечали на стимуляцию повышением частоты импульсации или коротким разрядом не менее чем в 50% проб.
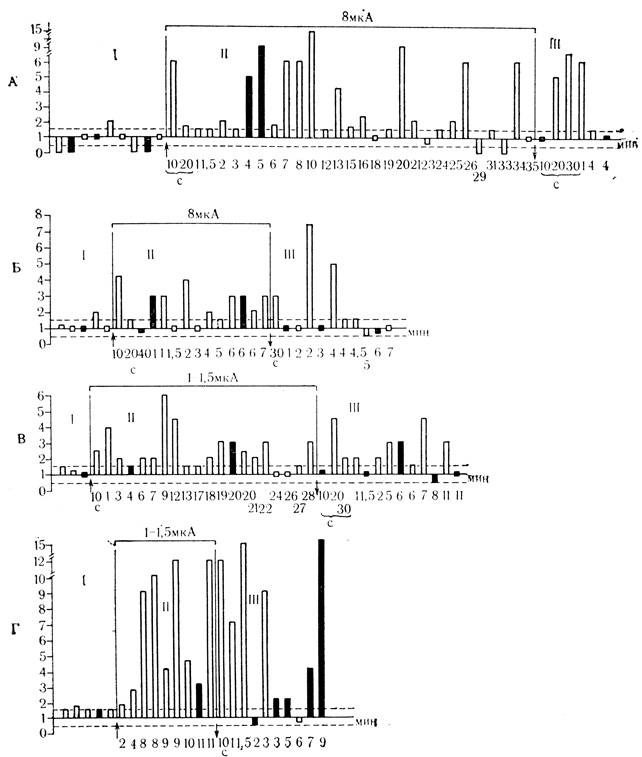
Рис. 38. Динамика ответов клеток двигательной коры на 'положительный' и 'отрицательный' звуковые стимулы на фоне суммарной поляризации и после выключения поляризующего тока. А - клетка на глубине 370 мкм. Б - клетка на глубине 1600 мкм. В - клетка на глубине 720 мкм. Поляризующий ток 1,5 мкЛ включен на 17-й минуте после начала поляризации током 1 мкА. Г - клетка на глубине 1560 мкм. В последействии - парадоксальная реакция на 'отрицательный' стимул. По оси ординат: отношение частоты разряда во время предъявления стимула (0,5 с) к среднему уровню фоновой активности, вычисленной за 2 с. до момента стимуляции. По оси абсцисс: время от момента включения до момента выключения поляризующего тока (стрелки). При спонтанной активности, равной нулю, по оси ординат откладывается величина, разная 2 при любой реакции клетки, и величина, равная 1 при отсутствии реакции. За уровень значимой реакции (пунктирная линия) принято изменение частоты разрядов на 50% по сравнению с фоновой. 'Положительный' стимул (не-штрихованные столбики) - тон 200 кол/с, 'отрицательный' (черные столбики) - тон 500 кол/с; I - ответы нейрона до поляризации; II - на фоне поляризации и III - после выключения поляризующего ток* (М. Я. Рабинович, Ф. В. Копытова, 1969)
Из 17 нейронов, в различной степени отвечавших на звуковые стимулы до поляризации, 6 клеток на фоне поляризации изменили характер реакции: у 3 клеток заметно усилился ответ на оба стимула в виде повышения частоты разрядов и сокращения их латентных периодов; 3 клетки, которые до поляризации отвечали на звук торможением активности, стали реагировать на оба стимула разрядом импульсов. Шесть нейронов до поляризации и на фоне поляризации отвечали только на один из двух предъявлявшихся стимулов, 5 нейронов, реагировавших на стимулы до поляризации, перестали на них отвечать во время поляризации. После выключения поляризующего тока соотношение клеток по характеру их реакций на оба стимула значительно изменилось, что можно видеть из данных табл. 9.
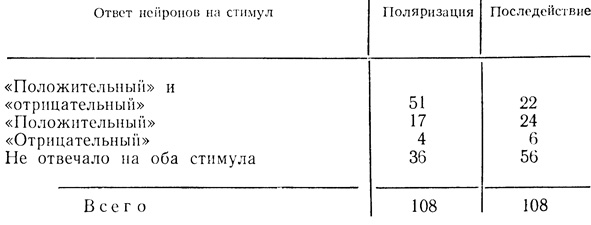
Таблица 9. Распределение нейронов двигательной коры по их ответам на звуковые стимулы 'положительного' и 'отрицательного' значения при поляризации и в последействии
Нас интересовали нейроны, которые на фоне поляризующего тока реагировали на оба звуковых раздражителя и затем в последействии отвечали только на стимул "положительного" значения. Таких клеток, как отмечалось выше, было 45 из числа не отвечавших ранее на звуковые стимулы и 6 клеток из числа изменивших характер ответов при поляризации. Следовательно, из 51 нейрона в последействии поляризации отвечали вспышкой разрядов только на "положительный" звуковой стимул 24 клетки. Таким образом, около 47% клеток проявили способность дифференцировать "положительный" раздражитель, многократно применявшийся на фоне поляризации, от "отрицательного", который предъявлялся на фоне поляризации не более 1 - 2 раз. Большинство других нейронов, как видно из табл. 9, реагировало равным образом на оба раздражителя или вовсе не отвечало ни на один из них.
На рис. 37 и 38 можно видеть, что нейроны, которые не реагировали в контрольных пробах на оба звуковых стимула или их ответы отличались незначительным повышением частоты разрядов, начинают четко отвечать на эти раздражители на фоне поляризации. После выключения поляризующего тока клетки на протяжении нескольких минут реагировали лишь на "положительный" стимул, который многократно применялся на фоне поляризации. На "отрицательный" стимул такие клетки не отвечали.
Представляют интерес те немногие нейроны, которые в последействии реагировали только на "отрицательный" стимул (рис. 38, Г). Таких клеток было 6, из них 3 клетки относились к группе из 45 нейронов, которые не отвечали на звуковые стимулы в контрольных пробах до поляризации, но реагировали на оба стимула при действии поляризующего тока. Эти клетки в последействии поляризации проявили парадоксальную реакцию: отвечали на стимул отрицательного значения и не реагировали или менее четко реагировали на "положительный" раздражитель.
Дифференцированные ответы на "положительный" и "отрицательный" звуковые стимулы в последействии поляризации часть нейронов сохраняла на протяжении 20-30 мин. Из 24 клеток, проявивших способность различать "положительный" и "отрицательный" раздражители, 20 нейронов сохраняли это свойство до 2 мин, 18 клеток - до 3 мин, 13 клеток - до 5 мин, 11 клеток - до 10 мин, 6 нейронов - до 20-30 мин. Один нейрон дифференцировал оба стимула на протяжении до 46 мин. Парадоксальные реакции также могли быть прослежены на протяжении до 10 мин. Через 30-35 мин после выключения поляризующего тока оба стимула вновь становились неэффективными.
Наиболее типичные примеры динамики ответов клеток на оба стимула при действии поляризующего тока и после его выключения представлены на графиках рис. 38. На фоне поляризации реакции на многократно применяемый "положительный" стимул не однозначны по интенсивности и подвержены колебаниям от пробы к пробе. Однако большинство реакций превышает уровень, принятый за значимый. В последействии поляризации клетки, как правило, некоторое время отчетливо дифференцируют стимулы "положительного" и "отрицательного" значения, а затем начинают отвечать на оба стимула. Ответы в последействии в одних клетках были примерно сходными по интенсивности разрядов с реакциями клетки на фоне поляризации (см. рис. 38, А, В), в других случаях - явно выше (см. рис. 38, Б, Г).
Обращает на себя внимание то, что при действии поляризации реакции клетки на "положительные" стимулы в некоторых пробах более выражены, чем на "отрицательные",- выше частота разрядов, короче латентный период. В этом феномене, по-видимому, проявляется способность нейронов к "разрабатыванию", т. е. повышению частоты разрядов и сокращению латентных периодов реакций при многократном повторении сенсорных стимулов (Л. Л. Воронин, В. Г. Скребицкий, 1967). Такое "разрабатывание" по отношению к несколько раз повторенному "отрицательному" стимулу в последействии поляризации можно видеть на рис. 38, Г. Однако большая часть клеток двигательной коры обычно не проявляла тенденции к "разрабатыванию" и их ответы на оба стимула при действии поляризации были сходными (см. рис. 38, А, Б, В).
Таким образом, основной результат исследования сводится к тому, что почти половина нейронов двигательной коры, недеятельных по отношению к примененным двум различным звуковым стимулам, начала реагировать на эти раздражители во время поляризации поверхности коры и в последействии отчетливо их различала в зависимости от предшествующего применения на фоне поляризации.
Уместно отметить, что количество клеток, проявивших способность различать сенсорный стимул в зависимости от предшествующих сочетаний, составляющее 47% всех исследованных клеток, соответствует среднему числу нейронов, у которых возникали условные реакции при различных формах выработки условного рефлекса и в различных корковых и подкорковых структурах мозга. Подобная аналогия может оказаться чисто внешней, но может и заключать в себе некоторые закономерности "сцепления" и взаимодействия на нейроне следов условной и безусловной стимуляции, что представляется нам более вероятным.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'