Зоны проекции сигнального стимула (слуховая кора и внутреннее коленчатое тело)
Слуховая кора. Условная активность нейронов слуховой коры изучалась на бодрствующих кроликах в экспериментальной обстановке, которая использовалась в исследовании условнорефлекторных реакций клеток двигательной коры.
Идентичными были и критерии верификации условных связей, однако акцент здесь ставится на воспроизведении активности пропущенного в отдельных сочетаниях подкрепляющего стимула (электрокожное раздражение конечности).
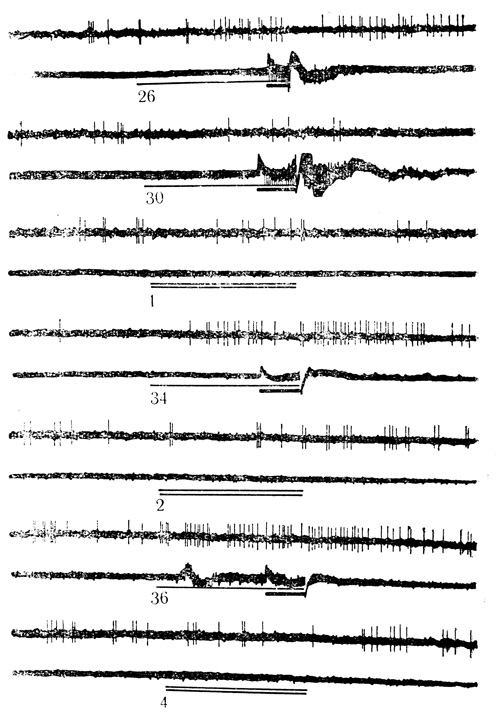
Рис. 31. Активирующие условные реакции нейрона слуховой коры на звук (щелчки 30 в 1 с) и торможение активности дифференцировочным стимулом (щелчки 10 в 1 с). Цифры на осциллограмме - число сочетаний или дифференцировок. Условный стимул обозначен тонкой линией, безусловный - толстой, дифференцировочный - двумя линиями. На каждой осциллограмме верхняя линия - разряды нейрона; нижняя линия - электромиограмма сгибателей передней конечности (Е. Л. Полонская, М. Я. Рабинович, 1971)
Всего зарегистрировано 115 нейронов (у 12 кроликов), из которых 51 клетка (около 44%) проявила условные реакции на звуковые стимулы (Е. Л. Полонская, М. Я. Рабинович, 1971). В группе условно реагирующих нейронов у 23 клеток (45%) ответы возникали в виде повышения частоты разрядов - такие нейроны проявляли активирующие условные реакции (рис. 31), 28 нейронов (55%) выработали условные ответы в форме торможения активности, т. е. проявили тормозные условные реакции (рис. 32). По сравнению с двигательной корой в слуховой корковой зоне значительно больше клеток (соответственно 42 и 55%) с тормозными условными реакциями, которые рассматриваются нами как одна из форм внутреннего торможения на клеточном уровне (М. Я. Рабинович, 1969, 1970).
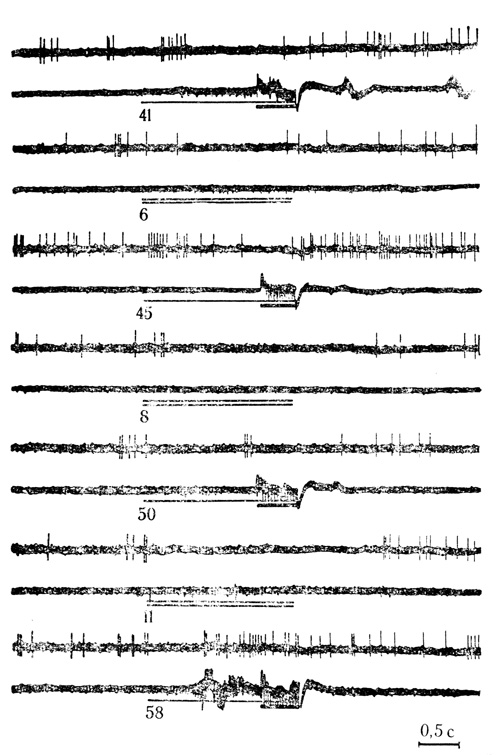
Рис. 31 Продолжение
Условные реакции нейронов, зарегистрированных первыми по ходу выработки двигательных условных рефлексов у каждого животного, возникали после 10-15 сочетаний и достигали значимого уровня после 20-25 сочетаний. Как правило, условные ответы нейрона опережали соответствующую двигательную реакцию животного (сгибание передней конечности), однако в отдельных сочетаниях клеточные реакции возникали после начала двигательного акта.
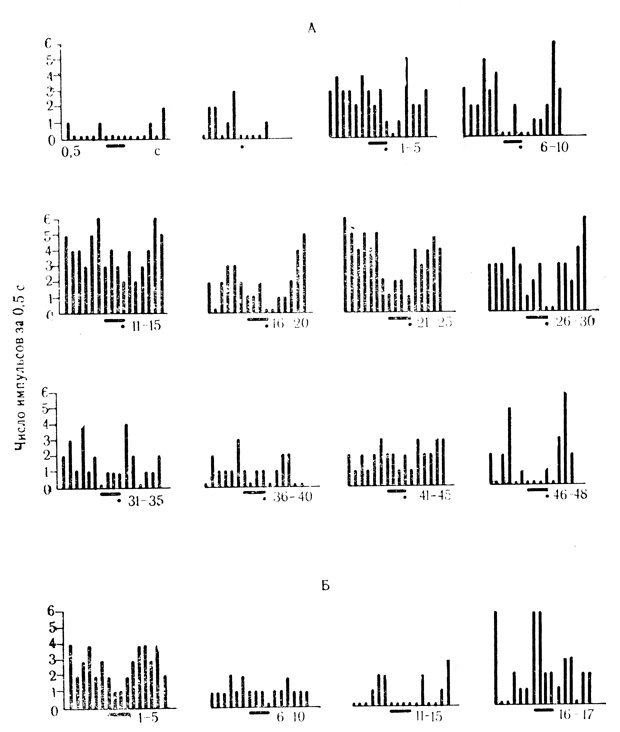
Рис. 32. Тормозные условные реакции нейрона слуховой коры на звук (щелчки 20 в 1 с) и растормаживание активности дифференцировочным стимулом (щелчки 10 в 1 с). Первые две гистограммы - контрольные пробы на звуковой стимул (черта) и электрокожное раздражение конечности (точка). Сочетаниям (А) и дифференцировкам (Б) на клетке (усреднены по 5 пробам) предшествовали 202 сочетания и 27 предъявлений дифференцировочного стимула па животчом (Е. Л. Полонская, М. Я. Рабинович, 1971)
Как видно из табл. 6, около 30% нейронов (35 клеток) в контрольных пробах, которые предшествовали каждой серии сочетаний на клетке, реагировали только на звуковые стимулы, около 18% клеток (21 нейрон) отвечали только на электрокожное раздражение, около 29% клеток (34 нейрона) проявили бимодальную активность, реагируя на оба стимула, около 23% клеток по отношению к сочетаемым стимулам оказались ареактивными. Надо иметь в виду, однако, что количество нейронов, реагирующих только на электрокожное раздражение, может быть несколько завышенным, так как контрольные пробы у большинства нейронов (кроме первых клеток у каждого животного) проводились после некоторого числа предшествовавших сочетаний, т. е. на фоне сенситизации клетки к действию подкрепляющего стимула.
Наиболее легко условные реакции вырабатывали нейроны, отвечающие в контрольных пробах на электрокожное раздражение. В группе нейронов, реагировавших до сочетаний только на этот стимул, условные реакции проявило около 62% клеток (13 из 21). В группе бимодальных клеток, отвечавших и на электрокожное раздражение, и на звуковые стимулы, условные реакции выработало около 59% нейронов (20 из 34). Из числа нейронов, до сочетаний реагировавших только на звук, условные реакции выработало 40% клеток (14 из 35). Четыре клетки, в группе не реагировавшие в контрольных пробах на оба стимула, по мере сочетаний проявили условные реакции активирующего и тормозного типа (см. рис. 32).

Таблица 6. Распределение нейронов, проявивших условные реакции на звук, в зависимости от характера предшествующих ответов на сочетаемые стимулы в контрольных пробах
Таким образом, формирование условной активности у нейронов корковой зоны проекции сигнального стимула связано преимущественно с приобретенной в процессе сочетаний способностью реагировать на подкрепляющий раздражитель. Вместе с тем очевидно, что в корковых структурах, куда адресуется условное раздражение, заметно возрастает роль сигнального фактора в формировании условных реакций клеток по сравнению с корковой зоной проекции подкрепления. В двигательной коре возбуждающая или тормозящая форма условной реакции нейрона определялась знаком ответа на подкрепление и только единичные клетки дали условные реакции по типу ответа на сигнальный стимул.
В слуховой коре из 14 нейронов, отвечавших в контрольных пробах лишь на звук и в последующем выработавших условные реакции, у 12 клеток условная активность протекала по типу ответов на звуковой стимул и только у 2 клеток наблюдались реакции противоположного знака - по характеру ответов на подкрепление. Из 13 нейронов группы, избирательно реагировавших в контрольных пробах на электрокожное раздражение, 10 нейронов проявили условные реакции по типу ответа на подкрепление, а 3 нейрона - противоположного знака, по типу реакции на звуковой стимул. Из 20 бимодальных нейронов у 15 клеток с однозначными ответами на оба стимула в контрольных пробах отмечалась условная активность, соответствовавшая знаку этих ответов; при разнозначных ответах 2 нейрона проявили условную активность по типу реакции на подкрепление, а 3 нейрона - по типу реакции на звуковой стимул.
Следовательно, более 1/3 клеток (18 из 51) слуховой коры выработало условные реакции в соответствии с активирующим или тормозящим действием сигнального стимула. В группе клеток, которая в контрольных пробах реагировала только на звук, большинство нейронов проявило условные реакции по знаку ответа на звуковые стимулы. В других группах по такому типу реагировала только небольшая часть клеток. Подкрепляющий фактор в данном случае стабилизирует, по-видимому, ответ клетки на звуковой стимул и придает нейрону свойства афферентной специализации, на что указывает способность таких клеток дифференцировать подкрепляемый и неподкрепляемый раздражители (см. рис. 31). Заслуживает быть отмеченным небольшое число нейронов, у которых по мере появления ответов на условный стимул изменялась реакция на подкрепляющее раздражение. У нейронов с активирующими условными реакциями (4 клетки) этот феномен проявляется в виде угнетения разрядов на подкрепление, доходящее до полного торможения ответа (рис. 33). У нейронов с тормозными условными ответами (4 клетки) угнетение разрядов на сигнальный стимул сопровождается растормаживанием реакции на подкрепляющее раздражение и повышением частоты разрядов по сравнению с ответом на изолированное действие безусловного стимула в контрольных пробах (см. рис. 20). Механизмы тормозящего и растормаживающего влияния условного ответа на подкрепление условнорефлекторной природы включаются в процессе сочетаний и выключаются при угашении условной реакции (см. рис. 20).
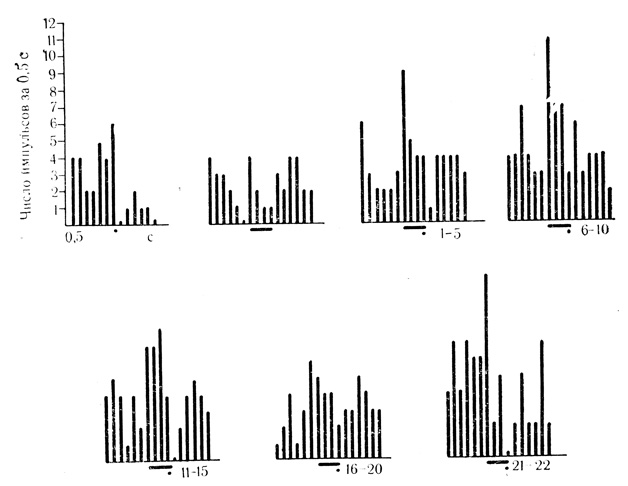
Рис. 33. Торможение ответов нейрона слуховой коры на подкрепляющий стимул по мере появления активирующих условных реакций на звук (щелчки 20 в 1 с). Первые две гистограммы - контрольные пробы на звуковой стимул Я (черта) и электрокожное раздражение конечности (точка). Под каждой гистограммой - число усредненных сочетаний для данной клетки, которым предшествовало 180 сочетаний на животном (Е. Л. Полонская, М. Я. Рабинович, 1971)
Фазирование условных реакций у таких клеток слуховой коры может и здесь служить указанием на их выходную функцию в популяции условнореагирующих нейронов структуры. Небольшое число таких клеток (около 16%, в том числе только 8% с активирующими реакциями), однако, свидетельствует о том, что они играют, по-видимому, второстепенную роль в передаче 1 условной активности на нейроны двигательной коры. Более вероятно предположить, что такие нейроны составляют специальный условнорефлекторный механизм регулирования элементами слуховой коры афферентного притока к подкорковым уровням слуховой системы.
Представляет интерес, что условнорефлекторные ответы большинства (54%) нейронов слуховой коры отличаются подчеркнуто волнообразной динамикой с чередованием периодов значимого уровня условной активности и полного прекращения ее без изменений обычных для клетки спонтанных разрядов. При этом обращает на себя внимание сравнительно небольшое число клеток (9 из 51), достигающих высокого уровня (80-100%) условных реакций. Условная активность остальных 42 нейронов не превышала 60-70% ожидаемых ответов от общего числа сочетаний. Особенностями динамики объясняется тот факт, что на фоне достаточно упроченного двигательного рефлекса значительная часть клеток отвечает на условный стимул так же, как при первых сочетаниях, и вырабатывает условные реакции как бы заново. В этой связи уместно отметить, что, по данным ряда авторов (Н. Н. Василевский, 1968; В. В. Шульговский, Б. И. Котляр, 1968; Miller e. a., 1972), важной особенностью реакций нейронов корковой проекции сигнального стимула является снижение интенсивности ответа по мере упрочения условного рефлекса. Достоверно уменьшается при этом и число условнореагирующих нейронов (Ю. А. Фадеев, 1969).
Эти наблюдения наряду с фактом преобладания в слуховой коре нейронов с тормозными условными реакциями (55%) и небольшим числом активирующихся клеток с предполагаемыми выходными функциями (8%) подтверждают давнее наблюдение М. Н. Ливанова, касающееся подвижной структуры афферентных звеньев дуги условного рефлекса. По-видимому, при реализации упроченного поведенческого акта действительно имеет место переключение по крайней мере части потока импульсов сигнальной стимуляции в сторону корковой зоны безусловного рефлекса на уровне подкорковых звеньев анализатора условного раздражения, минуя его корковые структуры (М. Н. Ливанов, 1952; М. Я. Рабинович, 1967; Т. С. Наумова, 1968; Н. Ю. Беленков, 1969).
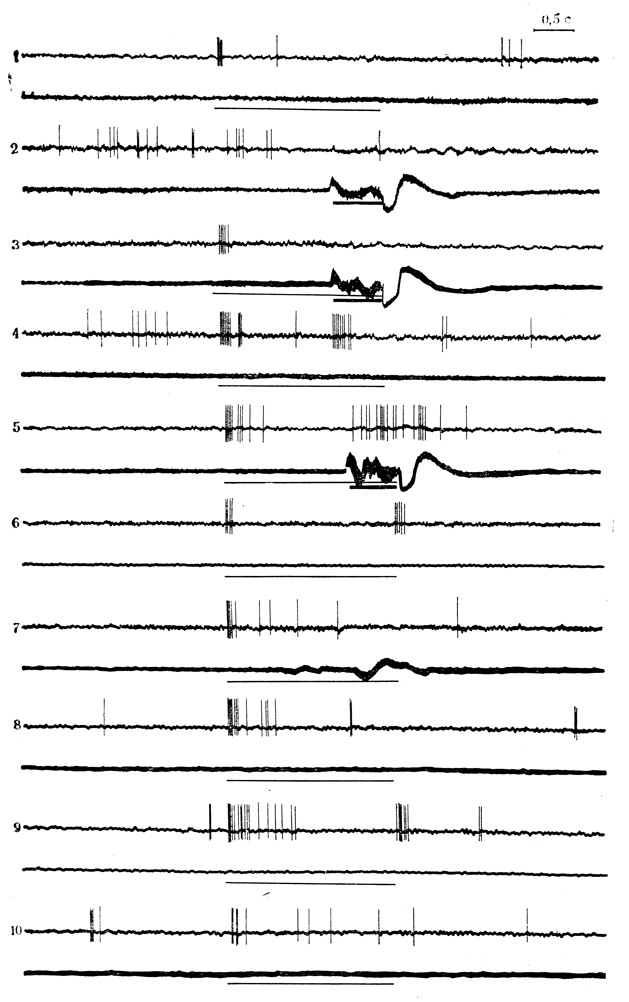
Рис. 34. Появление разрядов нейрона ВКТ на электрокожное раздражение конечности по мере сочетаний, воспроизведение их при пропусках подкрепления и угашение этой следовой реакции. 1, 2 - контрольные пробы на звуковой стимул (тонкая линия) и электрокожное раздражение (толстая линия); 3 - 8-е сочетание звука (щелчки 20 в 1 с) с подкреплением; 4 - пропуск подкрепления в 9-м сочетании; 5 - 21-е сочетание; 6 - 10 -соответственно 4, 15, 20, 25 и 35-я пробы на угашение. На каждой осциллограмме: верхняя кривая - разряды нейрона, нижняя - ЭМГ сгибателей передней конечности (Е. Л. Полонская, М. Я. Рабинович, 1972)
Внутреннее коленчатое тело (ВКТ). Условные реакции нейронов ВКТ исследовались в процессе выработки у бодрствующих кроликов двигательных условных рефлексов (сгибание передней конечности) сочетаниями звуковых стимулов (щелчки) с электрокожным раздражением контралатеральной конечности. У 13 подопытных животных зарегистрирована активность 82 нейронов мелкоклеточной части (pars principalis) ВКТ, верифицированных гистологически (на препаратах, окрашенных по Нисслю) по электрокоагуляционному пятну у кончика микроэлектрода (Е. Л. Полонская, М. Я. Рабинович, 1972).
По мере сочетаний достоверные условные реакции проявили 27 нейронов из 82 (около 33%). Из 27 условно реагирующих клеток 19 (около 70%) выработали активирующие условные реакции в виде повышения частоты разрядов в ответ на сигнальный стимул (рис. 34), у 8 клеток (около 30%) отмечалась тормозная условная активность в виде снижения частоты импульсации (рис. 35). У нейронов, зарегистрированных первыми в процессе сочетаний у каждого животного, условные реакции возникали после 10-15 сочетаний и достигали значимого уровня после 20-25 сочетаний.
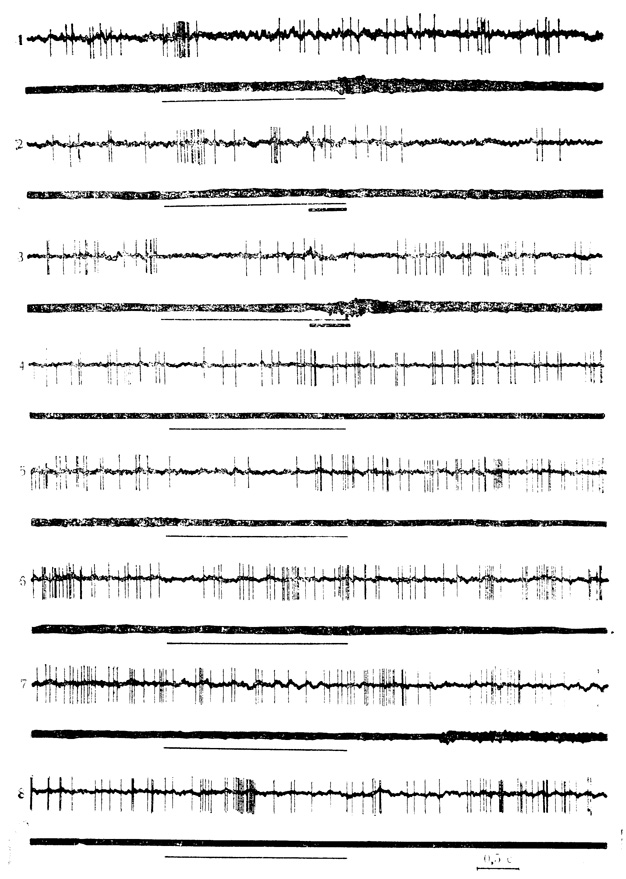
Рис. 35. Выработка тормозной условной реакции у нейрона ВКТ под влиянием угнетающего действия безусловного стимула и ее последующее угашение. 1 - контрольная проба на звуковой стимул (тонкая линия) 2 - 14-е (146-е), сочетание звука (щелчки 20 в 1 с) с электрокожным раздражением конечности (толстая линия); 3 - 26-е (158-е) сочетание; 4 - 8 - соответственно 5, 11, 16, 20 и 22-я пробы на угашение. Число сочетаний указано для данной клетки, в скобках - по отношению к животному. При угашении тормозной условной реакции клетки вновь появляется начальная пачка разрядов на звуковой стимул (Е. Л. Полонская, М. Я. Рабинович, 1972)
Критериями выработанной условной реакции клеток служили: появление ответов на условный стимул, который в контрольных пробах реакций не вызывал, или изменение по мере сочетаний знака первоначального ответа на щелчки (6 клеток); достоверное (более чем 50%) усиление ответов на условное раздражение с последующим их угашением (6 клеток); появление ответов на ранее недеятельное электрокожное раздражение конечности и воспроизведение следовой реакции на подкрепляющий стимул при контрольных пропусках его в сочетаниях с последующим угашением этой реакции (15 клеток). Пример сохранения ответа клетки на подкрепление при его пропусках и угашения следовой реакции на электрокожное раздражение представлен на рис. 34.
В контрольных пробах из 13 нейронов, зарегистрированных первыми в опыте у каждого из 13 животных, на щелчки реагировали 10 клеток; 9 клеток отвечали повышением частоты разрядов, одна клетка - торможением активности. 3 клетки на предъявленные звуковые стимулы не отвечали. На контрольные предъявления электрокожного раздражения конечности только один нейрон из 13 ответил весьма небольшой (1-2 спайки) и неустойчивой активацией, а другой - также слабо выраженным торможением. Обе клетки относились к числу реагировавших на щелчки. Однако по мере сочетаний некоторые нейроны приобрели способность реагировать на электрокожное раздражение конечности и в то же время прекратили отвечать на звуковые стимулы. Из табл. 7, в которой сведены данные о последующих контрольных реакциях на сочетаемые стимулы всех зарегистрированных нейронов, видно, что около 12% клеток (10 нейронов) стали отвечать только на электрокожное раздражение, потеряв способность реагировать на адекватные звуковые стимулы; около 24% клеток (20 нейронов) проявили бимодальную активность, реагируя на оба стимула, и только около 42% нейронов (34 клетки) оставались мономодальными и отвечали на одни звуковые стимулы.

Таблица 7. Распределение нейронов, проявивших условные реакции на щелчки, в зависимости от характера предшествующих ответов на сочетаемые стимулы
Приобретенная невосприимчивость к звуковым стимулам у клеток, начавших отвечать только на электрокожное раздражение, была достаточно устойчивой, так как из 10 таких нейронов лишь два выработали условные реакции на щелчки при последующих сочетаниях. По-видимому, в основе такого рода пластических сдвигов лежат какие-то специальные механизмы условнорефлекторной природы, избирательно блокирующие сигнал в системах вставочных нейронов данного афферентного канала. Возможно также, что эта функция реализуется кортико-фугальными влияниями выходных нейронов слуховой коры.
Из табл. 7 следует также, что наиболее легко условные реакции вырабатывали нейроны, у которых по мере сочетаний появились бимодальные реакции, чем и обеспечивалось постоянное соприкосновение на клеточном уровне сочетаемых стимулов и их взаимодействие. Но наличия бимодальной активности еще недостаточно для образования условной связи, так как из числа клеток с ответами на оба стимула условные реакции проявила только половина нейронов (10 из 20). В то же время 3 клетки из числа ареактивных в контрольных пробах при последующих сочетаниях выработали условные связи. По-видимому, для образования стабильной условной связи необходимо воздействие дополнительных факторов, которые привносятся подкрепляющим рефлексом, но нейрофизиологические механизмы которых остаются неясными. Структурной основой появления ответов на электрокожное раздражение у клеток основной части ВКТ могут служить межнейрональные связи этого образования с крупноклеточной частью ВКТ, на нейронах которого конвергируют как звуковые стимулы, так и раздражители сомато-сенсорных модальностей (Wepsic, 1966; Love, Scott, 1969; Raffaele e. a., 1969). 140
Следующие данные характеризуют особенности взаимодействия на нейронах ВКТ факторов сигнальной и подкрепляющей стимуляции, определяющих выработку активирующей или тормозной формы условной связи. В группе мономодальных нейронов, которые в контрольных пробах до сочетаний реагировали на щелчки, большинство клеток (10 из 12) выработало условные реакции в соответствии со знаком ответа на сигнальный стимул, 2 клетки - со знаком ответа на подкрепление; 2 нейрона, отвечавшие до сочетаний только на электрокожное раздражение, проявили условную активность по знаку реакции на подкрепляющий стимул. В группе из 10 нейронов, отнесенных в контрольных пробах к бимодальным, 4 клетки с однозначными реакциями на условный и безусловный стимулы выработали условные связи в соответствии со знаком этих реакций. При разнозначных ответах в этой группе условные реакции 2 клеток соответствовали знаку ответа на условный стимул, а 3 клеток - знаку ответа на подкрепление. Одна клетка с тормозными ответами в контрольных пробах на оба сочетаемых стимула выработала, однако, активирующую условную реакцию.
Таким образом, активирующая или тормозная форма условной реакции около половины нейронов ВКТ (12 из 27) формировалась под влиянием возбуждающего или угнетающего действия на клетку условного сигнала и примерно одна четверть нейронов (7 из 27) выработала условную реакцию в соответствии со знаком действия подкрепления. Примеры, иллюстрирующие выработку условной реакции по знаку действия на клетку сигнального и подкрепляющего стимулов, представлены на рис. 34 и 35 соответственно. В этой связи возрастает значение дополнительного приема идентификации условных реакций у нейронов, для которых сигнальный стимул является эффективным адекватным раздражителем с неизменяемым знаком ответа. Прием основан на появлении по мере сочетаний ответов клетки на ранее неэффективный безусловный стимул и сохранении следовой реакции на подкрепление при пропусках его в сочетании. Следовая реакция, приуроченная ко времени пропущенного подкрепления, угашается после ряда предъявлений неподкрепляемого сигнального стимула и восстанавливается при повторных сочетаниях, т. е. проявляет классические признаки условной связи (см. рис. 34).
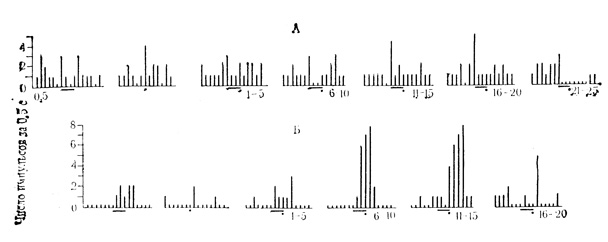
Рис. 36. Торможение и растормаживание ответов клеток ВКТ на подкрепляющий стимул по мере появления условных реакций. А - угнетение ответов на подкрепление у нейрона с активирующими условными реакциями. Сочетаниям на данной клетке (число под каждой гистограммой) предшествовало 19 сочетаний на животном; Б - усиление ответов на подкрепление у нейрона с тормозными условными реакциями. Сочетаниям на данной клетке предшествовало 160 сочетаний на животном. Первые две гистограммы на А и Б - контрольные пробы на звуковые стимулы и электрокожное раздражение конечности. Условный стимул - черта, подкрепление - точка (Е. Л. Полонская, М. Я. Рабинович, 1972)
Как и в других корковых и подкорковых структурах мозга, в ВКТ обнаружены нейроны, у которых при появлении ответов на сигнальный стимул изменялась реакция на подкрепление. У клеток с активирующими условными реакциями (4 нейрона) ответ на подкрепляющий стимул по мере сочетаний постепенно уменьшался и затем совершенно исчезал (рис. 36, А). У клеток с тормозными условными реакциями (4 нейрона) угнетение разрядов на сигнальный стимул сопровождалось выраженным растормаживанием реакции на подкрепление (см. рис. 36, Б).
Результаты исследования свидетельствуют о том, что нейроны основной (мелкоклеточной) части ВКТ способны к сложным перестройкам пластических свойств по мере сочетаний звуковых стимулов с электрокожными подкреплениями, которые у 33% клеток завершились выработкой условных реакций. Это, однако, намного меньше числа условнореагирующих нейронов, выявленных в корковых зонах проекции звукового сигнального стимула и подкрепляющего рефлекса, где в идентичной обстановке эксперимента условную активность проявили соответственно 44 и 52% клеток. Следует отметить также, что условные связи большинства нейронов ВКТ носили активирующий характер и только около 30% клеток выработали тормозные условные реакции.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'