Зона проекции подкрепляющего стимула (двигательная кора)
Исследования условной активности нейронов двигательной коры как зоны подкрепляющего стимула представляют особый интерес по ряду причин. Во-первых, хорошо очерченное сомато-топическое представительство в этой корковой зоне мышечных компонентов двигательного акта делает ее уникальным объектом для анализа динамики пластических свойств нейронов в процессе выработки локальных движений и корреляций между активностью клеток и поведенческими реакциями (Evarts, 1966; Woody e. a., 1970; Wood, Engel, 1972; Fetz, Finocchio, 1971, 1972). Во-вторых, нейроны моторной коры являются тем субстратом, в котором сложившаяся ассоциация между стимулами с различной сенсорной и биологической модальностями получает возможность выхода на эффекторные звенья условного двигательного поведения (П. К. Анохин, 1968).
Мы изучали условную активность нейронов моторной коры в процессе выработки у бодрствующих кроликов двигательных условных рефлексов сочетаниями звуковых стимулов (щелчки, тоны 200-500 кол/с) с электрокожным раздражением передней конечности. Зона коркового представительства соответствующей конечности предварительно определялась по двигательной реакции (сгибание конечности) на электрическое раздражение через истонченную кость черепа. Интервалы между сочетаниями варьировали в пределах от 40 до 90 с (Е. Л. Полонская, 1967; М. Я. Рабинович, 1970; Rabinovich, Polonskaya, 1973).
Каждое животное находилось в эксперименте 3-5 дней, в течение которых вырабатывался условный двигательный рефлекс на один и тот же звуковой стимул и регистрировалась активность отдельных клеток. Условные реакции части нейронов подвергались угашению. У другой части клеток вырабатывались дифференцировки на звуковой стимул другой частоты. Каждой серии сочетаний по отношению к отдельной клетке предшествовали контрольные пробы на звуковые стимулы (обычно той же интенсивности, но другой частоты, чем условные) и электрокожное раздражение конечности. Каждому животному предъявлялось от 200 до 750 сочетаний. Условная активность большинства нейронов была зарегистрирована на протяжении 40-70 сочетаний, а в отдельных случаях - до 100 сочетаний.
Всего исследовано 229 нейронов у 22 кроликов. Из этого числа клеток 118 нейронов (51,5%) проявили значимые условные реакции на звуковой стимул. К нейронам со значимыми условными реакциями относились клетки, отчетливо реагировавшие на условное раздражение на менее чем в 50% сочетаний. Критериями выработанной условной реакции на уровне нейрона служили появление стабильных ответов на условный стимул, который в контрольных пробах до сочетаний реакции не вызывал, изменение знака реакции в результате сочетаний, угашение условных ответов, относительно быстрое восстановление угашенных реакций новыми сочетаниями и выработка дифференцировок. Выработанная условная связь на уровне нейрона дополнительно идентифицировалась сохранением реакции клетки на месте, приуроченном во времени к однократно пропущенному в сочетании подкреплению.
Как следует из табл. 5, около 47% нейронов (108 клеток) в контрольных пробах до сочетаний реагировало только на электрокожное раздражение конечности, около 11 % нейронов (25 клеток) - только на звуковые стимулы, 34% нейронов (78 клеток) проявили бимодальную активность, реагируя на электрокожное раздражение и звуковые стимулы, и 8% нейронов (18 клеток) ни на один из этих стимулов не отвечали.

Таблица 5. Распределение нейронов, проявивших условные реакции на звук, в зависимости от характера предшествующих ответов на сочетаемые стимулы
Надо иметь в виду при этом, что мы стремились, особенно в начале исследования, отбирать нейроны, которые не отвечали на звук, но реагировали на электрокожное раздражение, с тем чтобы выработанные ответы могли быть без сомнений отнесены к условным. Это обстоятельство, по-видимому, несколько уменьшало число нейронов, отвечавших в контрольных пробах на звуковые стимулы.
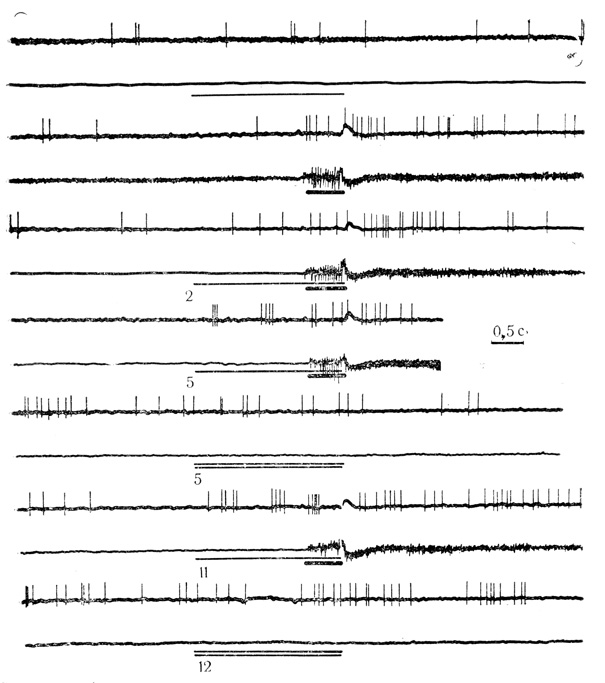
Рис. 27. Активирующие реакции нейрона двигательной коры на условный стимул (тон 200 Гц) и торможение активности дифференцировочным стимулом (тон 500 Гц). Клетка на глубине 1510 мкм. Условный стимул обозначен тонкой линией, безусловный - толстой, дифференцировочный - двумя линиями. Две верхние осциллограммы - контрольные пробы на звуковой стимул и электрокожное раздражение конечности. Цифры на каждой осциллограмме - число сочетаний или дифференцировок для данной клетки, которым предшествовали 134 сочетания и 50 проб на дифференцировку на животном. На каждой осциллограмме: верхняя линия - разряды нейрона; нижняя- электромиограмма сгибателей передней конечности (М. Я. Рабинович, Е. Л. Полонская, 1973)
Из 118 нейронов, выработавших условные реакции на звуковые стимулы, у 68 клеток (около 58%) ответы возникали в виде повышения частоты разрядов (активирующие условные реакции); 50 клеток (около 42%) проявили условные ответы в форме торможения фоновой активности (тормозные условные реакции). Примеры активирующих и тормозных условных ответов представлены на рис. 27 и 28.
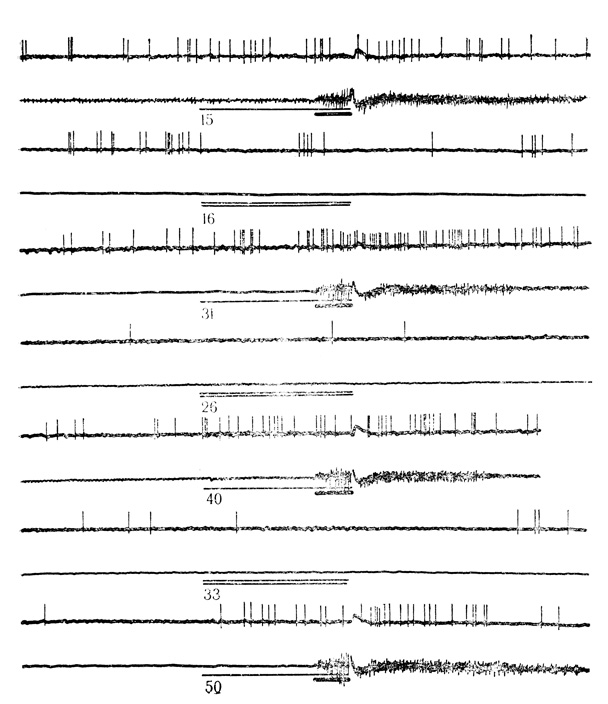
Рис. 27. Продолжение
Наиболее легко условные реакции вырабатывали нейроны, проявившие способность к полисенсорной активности. Из табл. 5 видно, что около 78% нейронов с бимодальными реакциями выработало условные ответы на звуковые стимулы (61 клетка из 78); из группы клеток, реагировавших в контрольных пробах только на звуковые раздражители, условные реакции проявили 56% нейронов (14 клеток из 25); из группы нейронов, до сочетаний, отвечающих только на электрокожное раздражение, условную активность проявили около 34% клеток (37 нейронов из 108); 6 нейронов (около 33%) из числа не реагировавших в контрольных пробах на оба стимула, по мере сочетаний выработали условные реакции активирующего или тормозного типа.
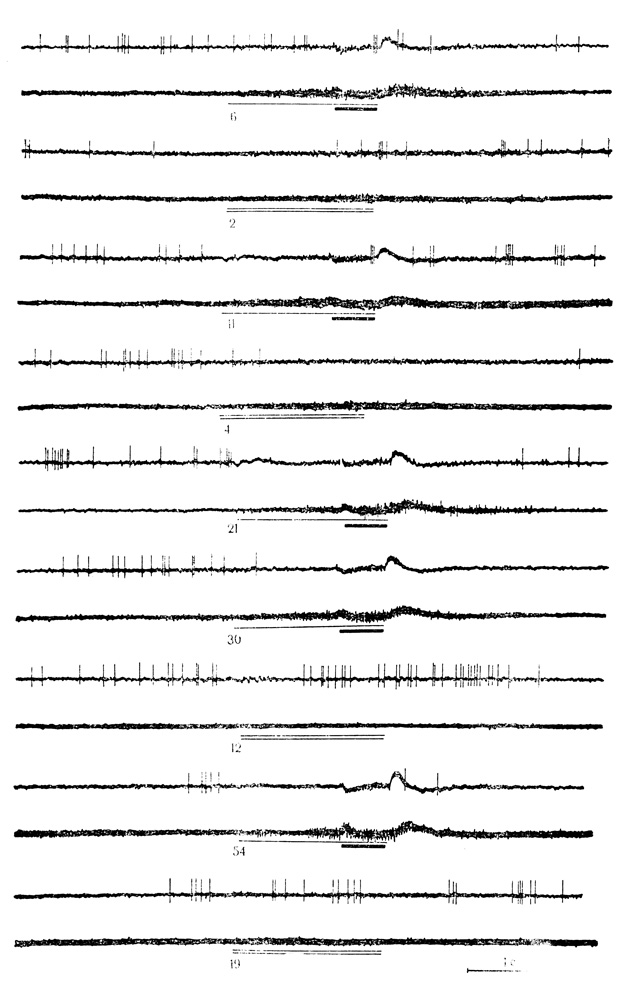
Рис. 28. Тормозные реакции нейрона двигательной коры на условный стимул (щелчки 20 в 1 с) и растормаживание активности дифференцировочным стимулом (щелчки 10 в 1 с). Клетка на глубине 1400 мкм. Сочетаниям и дифференцировкам на клетке предшествовали 444 сочетания и Доб предъявлений дифферепцировочного стимула на животном. Другие обозначения те же, что на рис. 27 (М. Я. Рабинович, Е. Л. Полонская, 1973)
Необходимо подчеркнуть, что эти соотношения следует рассматривать только как исходные, характеризующие относительно ранние этапы выработки условной связи. По мере упрочения и стабилизации условного рефлекса у животного пластические свойства клеток претерпевают эволюцию, после примерно 400 сочетаний элементы с моносенсорными реакциями только на электрокожное раздражение конечности не обнаруживаются и все исследованные нейроны из числа выработавших условные реакции в контрольных пробах отвечают на оба стимула, проявляя бимодальную активность (рис. 29).
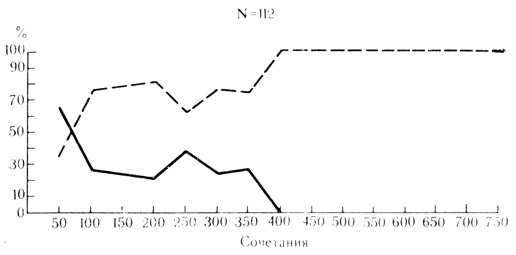
Рис. 29. Эволюция пластических свойств нейронов двигательной коры по мере выработки условных рефлексов у животного. Сплошная линия - моносенсорные клетки, реагировавшие в контрольных пробах только на электрокожное раздражение конечности; пунктирная линия - бимодальные нейроны, отвечавшие в контрольных пробах на оба стимула - звуковой и электрокожный. По оси абсцисс - число сочетаний; по оси ординат - процент нейронов, проявивших указанные реакции (М. Я. Рабинович, Е. Л. Полонская, 1973)
По данным Olds и Hirano (1969), полученным на крысах, число нейронов, усиливших ответы на звуковой стимул или начавших реагировать на него в процессе сочетаний с пищевым подкреплением, увеличилось в гиппокампе и неспецифических структурах таламуса в 5 раз по сравнению с контрольной группой, а в гипоталамусе и преоптической области - в 2 раза. В. Б. Швырков (1969) показал, что в сомато-сенсорной зоне коры кроликов до сочетаний с оборонительным подкреплением на свет реагировало 27%, а после сочетаний - 47% клеток. Однако в той же сомато-сенсорной коре кроликов В. Б. Швырковым (1970) были обнаружены клетки, на которые проецировалось электрокожное раздражение конечности, но не проявивших вместе с тем условных реакций на вспышки света, синхронизированные со звуковыми щелчками, т. е. так и не приобретших способности отвечать на эти сенсорные стимулы.
Неожиданным оказался относительно небольшой процент условнореагирующих нейронов из числа отвечавших в контрольных пробах только на электрокожное раздражение: таких клеток было всего 34%. По-видимому, здесь получили отражение процессы концентрации условной активности по мере сочетаний в ограниченной популяции нейронов, непосредственно связанной с соматотопической проекцией мышечных компонентов выработанного двигательного акта и локальными влияниями при этом обратной проприоцептивной импульсации на пластические свойства этих клеток (Л. Л. Воронин и др., 1969; М. Я. Рабинович, 1970; Woody е. а." 1970; Fetz, Finocchio, 1972; Woody, Engel, 1972). Возможно, что стабильные условные реакции обнаруживают преимущественно нейроны корковой зоны локализации подкрепляющего двигательного акта, тогда как ответы на электрокожное раздражение вне сочетаний способны проявить клетки в значительно более широких пределах проекционной зоны этого стимула.
Условные разряды нейронов, зарегистрированных первыми в процессе выработки у данного животного условного рефлекса, возникали после 5-15 сочетаний и достигали значимого уровня после 20-30 сочетаний, т. е. задолго до выработки условного движения конечности, которое в обстановке нашего эксперимента появлялось после 150-200 сочетаний.
Как отмечалось выше, на один и тот же положительный условный стимул нейроны способны были выработать как активизирующие, так и тормозные условные связи. Эти две основные формы условных ответов в большинстве случаев определялись возбуждающим или угнетающим действием на клетку подкрепления независимо от знака реакции ее на звуковой стимул в контрольных пробах до сочетаний. В некоторых случаях влияние подкрепляющего стимула на знак условного ответа клетки сказывалось также, если реакция на него наступала не в момент предъявления, а в последействии. Только изредка встречались клетки, которые вначале реагировали на подкрепление торможением разрядов, но по мере сочетаний вырабатывали активирующие условные связи по знаку действия на нейрон сигнального стимула.
По данным O'Brien и Fox (1969a), перестройку условных реакций по знаку подкрепляющего действия проявило 79% нейронов двигательной коры кошек, у которых условные рефлексы вырабатывались сочетаниями света с электрокожным раздражением конечности. Остальные нейроны реагировали на условный и безусловный стимулы активностью с противоположным знаком. По-видимому (если судить по материалам обсуждения результатов), к числу таких клеток авторы относили нейроны, у которых условная активность по мере сочетаний вызывала торможение (или растормаживание) ответов на подкрепление. O'Brien и Fox такие нейроны не выделяли. 126
Между тем клетки, у которых по мере выработки и упрочения ответов на условный стимул изменялась реакция на подкрепляющее раздражение, представляют значительный интерес (см. главу III, с. 85). Таких нейронов в моторной коре обнаружено около 25% (29 из 118). У нейронов с активирующими условными реакциями (18 клеток) этот феномен проявлялся в виде постепенного угнетения разрядов на подкрепление, доходящее часто до полного торможения ответа (см. рис. 19). У нейронов с тормозными ответами (11 клеток) угнетение разрядов на условный стимул сопровождалось растормаживанием реакции на подкрепляющее раздражение и часто значительным повышением частоты разрядов па сравнению с ответом на изолированное действие безусловного стимула в контрольных пробах (рис. 30). Эффекты тормозящего и растормаживающего влияний условного ответа на подкрепление были тем лучше выражены, чем интенсивнее проявлялась условная реакция,, сохранялись некоторое время в последействии сочетания, и постепенно исчезали при угашении.
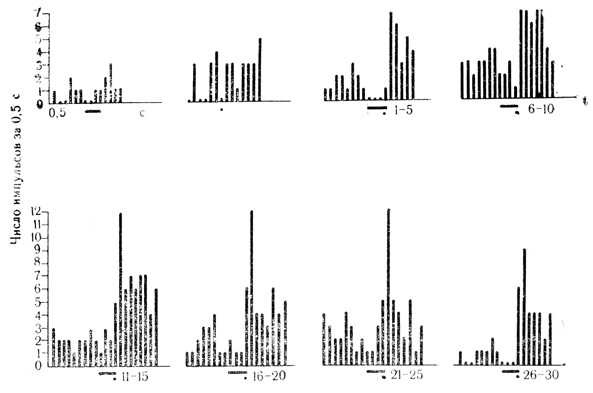
Рис. 30. Растормаживание ответов нейрона двигательной коры на подкрепляющий стимул по мере появления тормозных условных реакций на щелчки 20 в 1 с. Клетка на глубине 1050 мкм. Под каждой гистограммой - число сочетаний Для данной клетки, которым предшествовало 258 сочетаний на животном. Первые две гистограммы - контрольные пробы на звуковой стимул (черта и электрокожное радражение конечности (точка) (М. Я. Рабинович, Е. Л. Полонская, 1973)
Отчетливая фазность реакций на условный и безусловный стимулы, приобретаемая в процессе сочетаний, позволяет предположить, что эти нейроны моторной коры осуществляют конечную передачу двигательного сигнала на спинальные уровни рефлекса и являются, следовательно, выходными клетками в цепи условно реагирующих нейронов. В пользу такого предположения свидетельствуют наблюдения Evarts (1966). По его данным, идентифицированные нейроны пирамидного тракта, реализующие передачу условного двигательного сигнала по быстропроводящим аксонам, отличаются от других клеток моторной коры относительно кратковременным, фазным повышением частоты разряда, которое предшествует движению. Наблюдение Evarts согласуется с данными П. Г. Костюка (1971) о том, что на спинальном уровне существует специализированная система вставочных нейронов, приспособленных к выделению из общей нисходящей испульсации фазных компонентов двигательного сигнала для последующей передачи их на соответствующие мотонейроны.
| Https://barbellsport.ru/ barbellsport.ru |
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'