Время поведенческой реакции и латентные периоды условных ответов клеток
Сопоставления активности единичных клеток со сложными, часто многокомпетентными, поведенческими актами встречают пока значительные трудности. Это видно на примерах разнообразных корреляций, проведенных в ряде интересно задуманных и хорошо выполненных исследований (Н. Н. Василевский и др., 1972, б, в; Fetz, 1969; Woody e. a., 1970; Fetz, Finocchio, 1971, 1972; Woody, Engel, 1972). Вместе с тем такой анализ необходим, и первым приближением к нему могут служить измерения временных соотношений между условной активностью нейронов и относительно простыми условными актами животного.
Первые наблюдения в этом аспекте принадлежат Evarts (1966), который сопоставил латентные периоды условных разрядов идентифицированных корковых нейронов пирамидного тракта со временем специальной двигательной реакции (разгибание кисти) в процессе выработки у обезьян пищедобывательных условных рефлексов. Оказалось, что вспышка разрядов у таких клеток возникает через 100-120 мс после начала действия сигнального стимула (свет), а условная двигательная реакция появляется только через 200-220 мс, т. е. в среднем на 100 мс позже. Таким образом, время условной поведенческой реакции примерно в 5 раз превышает время, которое необходимо для проведения импульсов от "корковых клеток к исполняющим команды мышцам и обосновано структурно-функциональной организацией пирамидного и экстрапирамидного трактов (П. Г. Костюк, 1967; Phillips, 1967).
При этом важно, что величины латентных периодов электромиограммы разгибателей кисти и самого движения отчетливо коррелируют с латентностью предшествующего условного разряда клетки - наиболее короткие латентные периоды разряда клетки соответствуют минимальным латентным периодам поведенческого акта. Это может означать, что удлиненное время условной двигательной реакции - не случайное явление, а отражение каких-то неизвестных закономерностей, которые возникают при становлении условной связи и характеризуют ее.
По данным Kubota и Niki (1971), Kubota с соавторами (1972), Niki с соавторами (1972), активирующие условные реакции нейронов префронтальной коры обезьян на зрительные стимулы возникали с латентными периодами до 300 мс и опережали появление отсроченной пищедобывательной реакции на 300-400 мс. Нейроны хвостатого ядра, дорсо-медиального и вентро-латерального ядер таламуса реагировали на сигнальный стимул с латентными периодами в пределах 150 мс, опережая двигательную пищедобывательную реакцию в среднем на 200 мс.
Fetz и Finocchio (1971, 1972) при выработке у обезьян инструментальных пищедобывательных условных рефлексов обнаружили, что максимум разрядов клеток моторной коры опережает двигательную реакцию проецируемой на эти нейроны конечности на 700-100 мс, что соответствует измерениям Evarts (1966). Несколько меньшие значения приводят Luschei с соавторами (1971) при выработке у обезьян условнорефлекторного сжимания челюсти: активирующие и тормозные условные реакции нейронов корковой зоны проекции жевания (нижняя треть прецентральной извилины) опережали сокращение жевательных мышц в среднем на 50 мс со стандартным отклонением около 20 мс.
Об относительно большом разрыве клеточных условных реакций и соответствующих поведенческих актов свидетельствуют также измерения латентных периодов условных разрядов гамма- и альфа-мотонейронов, проведенные Buchwald с соавторами (1961) у кошек с предварительно выработанными условными рефлексами на звуковые стимулы. Разряды волокон гамма-эфферентной системы возникают через 18 мс после начала действия сигнального раздражения и предшествуют разрядам альфа-волокон и условному сгибанию конечности, которые появляются всегда одновременно с латентным периодом 50-100 мс. Подсчеты показали, что эффекты звуковой стимуляции после соответствующих переключений могут достигать альфа-мотонейроны за 26 мс. Между тем даже минимальная латентность условного поведенческого акта превышает это время на 30 мс, а максимальная - на 80 мс.
Не менее интересные результаты дают измерения латентных периодов реакций нейронов на один и тот же сенсорный стимул до и после сочетаний. По подсчетам Evarts (1966), латентность разрядов клеток моторной зоны коры обезьян на условный стимул увеличилась в процессе сочетаний в среднем на 70 мс, происхождение которых он затрудняется объяснить.
Почти в 3 раза, по данным O'Brien, Fox (1969), возрастают латентные периоды максимальных условных реакций нейронов моторной коры кошек на световые раздражители по сравнению с контрольной стимуляцией. И. Н. Василевский (1968) связывает выработку подлинной условной активности клеток сомато-сенсорной коры кроликов с появлением ответов на сигнальные световые или звуковые стимулы с латентными периодами в пределах 500-1200 мс.
В экспериментах на бодрствующих кошках с хронически вживленными электродами Halas с соавторами (1970) показали, что при выработке оборонительных условных рефлексов на звук латентные периоды условных реакций популяций нейронов (multiple units) в коре и подкорковых звеньях слуховой системы достигают 500 мс после примерно 60 сочетании. Увеличение латентных периодов условных реакций на звук нейронов слуховой коры в процессе выработки у кошек условных мигательных рефлексов отмечает также У. Г. Гасанов (1972, а, б).
В нашей лаборатории Л. Л. Воронин и Р. Г. Кожедуб (1971, а) вырабатывали клеточные аналоги условного рефлекса на нейронах двигательной коры бодрствующих кроликов сочетаниями неэффективной ("условный" раздражитель) и эффективной ("безусловный" раздражитель) электростимуляции различных точек корковой поверхности. В такой упрощенной экспериментальной ситуации было обнаружено, что при предъявлениях сочетаний с короткими интервалами между ними (1-5 с) на ранее неэффективные стимулы появлялись крайне неустойчивые ответы, которые самопроизвольно исчезали без угашения и никогда не меняли знака реакции по знаку действия подкрепления. Подобного характера ответы на "условный" стимул возникали при пробах на "псевдообусловливание" (предъявления "условного" и "безусловного" стимулов вне сочетаний), что послужило основанием идентифицировать их как проявление неспецифической сенситизации. Латентные периоды таких реакций не превышали 20-30 мс (рис. 23, А).
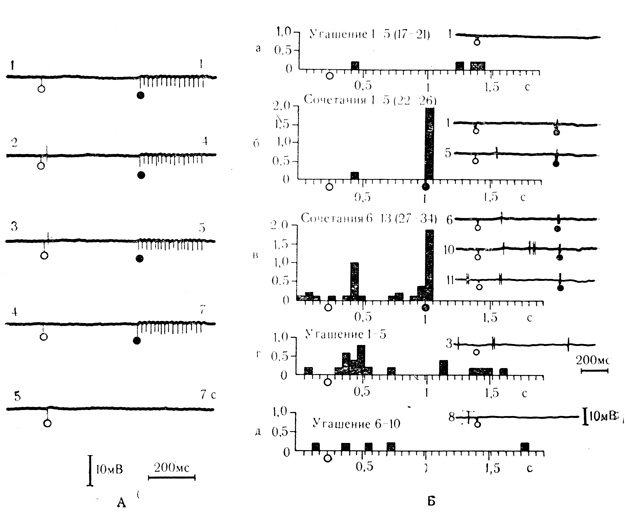
Рис. 23. Латентные периоды ответов клеток двигательной коры на условный стимул при неспецифической сенситизации и при выработанной условной связи. А - неспецифическая сенситизация. Реакция на условный стимул исчезает после 5 сочетаний (цифры справа). Предъявление условного стимула через 7 с после последнего сочетания неэффективно (осциллограмма 5); Б - клеточный аналог условного рефлекса, б и в - сочетания; а, г и д - угашение реакции. Цифры над гистограммами и слева от осциллограмм - порядковые номера сочетаний для данной клетки (в скобках для всего опыта). По оси ординат - среднее число импульсов на интервал анализа: по оси абсцисс - разметка интервалов анализа и время. Белые кружки - предъявление условного стимула, черные кружки - подкрепление (Л. Л. Воронин, Р. Г. Кожедуб 1971, а)
При сочетаниях с более длительными межсигнальными интервалами (12-120 с) около 43% клеток выработало подлинные активирующие или тормозные условные реакции в соответствии с возбуждающим или угнетающим действием на клетку подкрепляющего стимула. Выработанные реакции угашались при отмене подкрепления и быстро восстанавливались при повторных сочетаниях; при пропусках подкрепляющего стимула в сочетании на его месте возникала следовая реакция, воспроизводившая действие подкрепления. Латентные периоды ответов возрастали при этом до 150-200 мс (рис. 23, Б).
Существуют и противоположные изложенным выше результаты анализа динамики латентных периодов условных реакций клеток в процессе выработки условных рефлексов. Olds с соавторами (1972) вырабатывали у крыс пищедобывательные условные рефлексы на звуковой стимул (тон) с фиксированным интервалом предъявления сочетаний (около 1 мин). Авторы ставили перед собой задачу выявить "первично обучающиеся" нейроны, к числу которых относили клетки, реагирующие на условный стимул с латентными периодами в пределах первых 20 мс. Клетки, которые отвечали на сигнальный стимул в пределах до 80 мс, были классифицированы как вторично реагирующие, отражающие уже где-то выработанную условную связь. Нейроны, которые отвечали на условный сигнал в пределах остального времени его односекундного изолированного действия, исключались из анализа.
В соответствии с принятыми критериями оценки условных реакций нейронов наибольшее число "первично обучающихся" клеток было обнаружено в заднем ядре таламуса (53%), а наименьшее - в сенсо-моторной коре (3%), во фронтальной коре (4%), в поле СА-3 гиппокампа (5%), внутреннем коленчатом теле (14%) и т. п. По этому же критерию ряд образований мозга (поле СА-1 гиппокампа, наружное коленчатое тело, вентральные ядра таламуса, тектум, передне-дорсальная часть ретикулярной формации среднего мозга) были отнесены к структурам, нейроны которых не способны к обучению. Отсюда авторы делают далеко идущее заключение, что "первично обучающиеся" нейроны строго локализованы в определенных структурах мозга на всех его уровнях и что между зонами мозга способными и не способными к обучению существуют жесткие и неизменные границы.
Расхождения между данными большинства исследователей и результатами Olds с соавторами (1972) могут быть объяснены принципиально важными упущениями в анализе и трактовке этими авторами экспериментальных фактов. Прежде всего это относится к процедуре выработки пищедобывательных условных рефлексов. Применив фиксированный межсигнальный интервал (около 1 мин), исследователи неизбежно вырабатывали условную реакцию клетки на время. По данным нашей лаборатории (Ф. В. Копытова, М. Я. Рабинович, 1967; Ф. В. Копытова, Л. К. Куликова, 1970, 1971; Ф. К. Копытова, И. Н. Боголепова, 1972; Ф. В. Копытова, Ю. С. Медникова, 1972; М. Я. Рабинович, 1971), условная реакция нейронов на время в различных структурах мозга (двигательная кора, гиппокамп, миндалина) проявляет себя в значительном числе проб опережающим возбуждением, которое предшествует с вариабельными временными параметрами предъявлению очередных сочетаний (см. рис. 45, 47, 49, 50).
При усреднениях такого рода ответов, как это делали авторы, в результате будут иметь место как коротколатентные реакции, в пределах до 20 мс, так и нулевые реакции, поскольку опережающие сигнальный стимул разряды клетки могут быть приняты за межсигнальные реакции.
Как следует из приведенной авторами сводной таблицы, большинство нейронов в каждом из исследованных образований мозга реагировало на сигнальный стимул с латентными периодами в пределах до 80 мс, в том числе и в заднем ядре таламуса - наиболее "обучаемой" структуре мозга. В этом ядре с латентными периодами до 20 мс реагировало 53% клеток, а с латентными периодами до 80 мс - 88% клеток (при подсчетах по отношению к общему числу исследованных нейронов образования, отвечавших в интервале от 0 до 80 мс). В сенсо-моторной коре с латентными периодами до 20 мс реагировало только 3% клеток, а в пределах до 80 мс отвечало 22% клеток; 25 нейронов этой структуры (из 32 исследованных клеток) было исключено из анализа. В структурах, которые были отнесены к числу "необучаемых", например, в поле СА-1 гиппокампа, на сигнальный стимул с латентными периодами до 80 мс реагировало 20% клеток; 29 нейронов из 36, зарегистрированных в этом поле, были исключены из анализа. Приняв явно искусственный, экспериментально и теоретически необоснованный критерий отбора условнореагирующих нейронов, авторы исключили, таким образом, из анализа 61% клеток, значительная часть которых отвечала на условный стимул в пределах, превышающих начальные 80 мс его действия.
Следует отметить, однако, что в одной из предшествующих работ (Olds, Hirano, 1969), когда при выработке у крыс пищедобывательных условных рефлексов на звук интервалы между сочетаниями варьировали (сочетания предъявлялись "по случайному временному расписанию"), результаты измерений латентных периодов условнореагирующих клеток были иными. В этой работе Olds и Hirano (1969) показали, что условная активность нейронов гиппокампа, латерального и медиального гипоталамуса, преоптической области и ретикулярной формации среднего мозга была наиболее отчетливо выражена на протяжении последних 500 мс односекундного изолированного действия сигнального раздражителя (поздний компонент условного ответа). Ранние компоненты условной реакции клеток ( в первые 300 мс действия условного стимула) были выражены заметно менее отчетливо. По-видимому, результаты данного исследования ближе к истине и позволяют полагать, что повышение латентных периодов реакций клеток на стимулы, приобретшие сигнальное значение, представляет собой важную и характерную особенность условно-рефлекторного процесса, которая остается пока загадочной.
Использование фиксированных интервалов между сочетаниями сказалось также на результатах измерений латентных периодов условных реакций нейронов и поведенческого акта в исследовании Woody с соавторами (1970), которые вырабатывали у кошек мигательные условные рефлексы на звук. Вместе с тем даже в такой экспериментальной ситуации было обнаружено, что латентные периоды условного мигания возрастают в процессе сочетаний с 7 мс в среднем (контрольные пробы) до 20 мс. При этом опережающая поведенческий акт активация нейронов коронарно-прецентральной зоны коры возникает с вариабельными латентными периодами в пределах 8-16 мс.
При анализе временных соотношений между условными реакциями клеток и поведенческими актами животного следует иметь в виду закономерности "недеятельной" фазы условного рефлекса, которую И. П. Павлов, как известно, относил к разновидности внутреннего торможения - запаздыванию и длительность которой каким-то образом зависит от интервала отставления подкрепляющего стимула.
Однако предваряющая условную реакцию фаза торможения, в какой-то мере объясняя возможный генез увеличения латентных периодов выработанных рефлексов, не раскрывает природы довольно большого разрыва во времени между условными разрядами клеток и соответствующей условной реакцией животного. Напомним, что, по данным Evarts (1966), временные соотношения между условными разрядами клеток и поведенческими реакциями удивительно сохраняются при изменениях величин латентных периодов на уровне нейрона.
Дальнейшие исследования в этом направлении представляются нам одной из первоочередных задач структурно-функционального анализа условного рефлекса на уровне микрореакций отдельных элементов. Возможно, что решение вопроса будет приближено, если обратить внимание на достоверную идентификацию природы и особых свойств нейромедиаторов синаптических входов для условного и безусловного стимулов, и входов для общего пути, передающего уже сложившуюся условную связь с одного нейрона на другие в целях клеточных популяций.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'