Торможение и растормаживание подкрепляющего действия
Для характеристики механизмов взаимодействия сигнального и подкрепляющего раздражителей на уровне нейрона представляют интерес клетки, у которых по мере появления ответов на условный стимул изменялась реакция на подкрепление. У нейронов с активирующими условными реакциями этот феномен проявляется в. виде угнетения разрядов на подкрепляющий стимул (рис. 19), часто до полного их торможения на фоне отчетливо выраженного движения конечности (М. Я. Рабинович, Е. Л. Полонская, 1969).
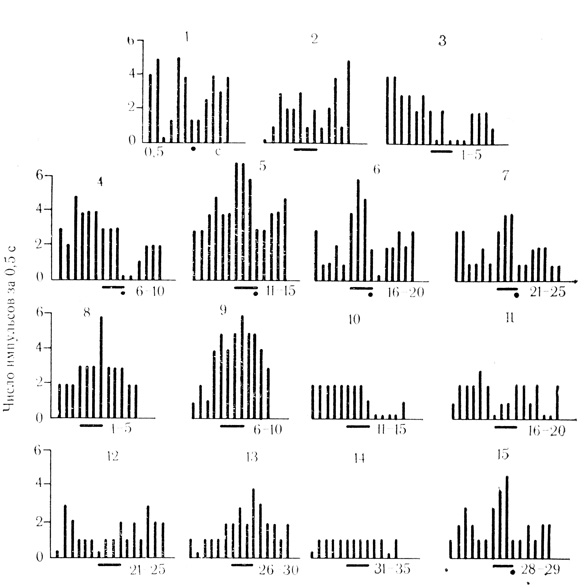
Рис. 19. Торможение ответов нейрона двигательной коры на подкрепляющий стимул по мере появления активирующих условных реакций на звук (щелчки - 20 в 1 с) и угашение тормозного действия. Под каждой гистограммой (3-7) усредненное число сочетаний или (8-14) число проб на угашение; 1, 2 - контрольные пробы на электрокожное раздражение конечности (точка) и звук (черта). Нейрон на контрольные стимулы не реагировал и был отнесен к ареактивным клеткам; 15 - восстановление условной реакции повторными сочетаниями
У нейронов с тормозными условными ответами снижение частоты разрядов на сигнальный стимул сопровождается растормаживанием реакции на подкрепляющее раздражение в виде значительного повышения частоты импульсации по сравнению с ответами на изолированное действие безусловного стимула в контрольных пробах (рис. 20). Торможение и растормаживание эффектов подкрепления проявили, по нашим данным, около 25% клеток двигательной зоны коры из числа выработавших условные реакции, около 16% клеток слуховой зоны коры и около 30% клеток внутреннего коленчатого тела - при выработке у бодрствующих кроликов двигательных условных рефлексов на звук с электрокожным подкреплением (Е. Л. Полонская, М. Я. Рабинович, 1971, 1972; Rabinovich, Polonskaya, 1973).
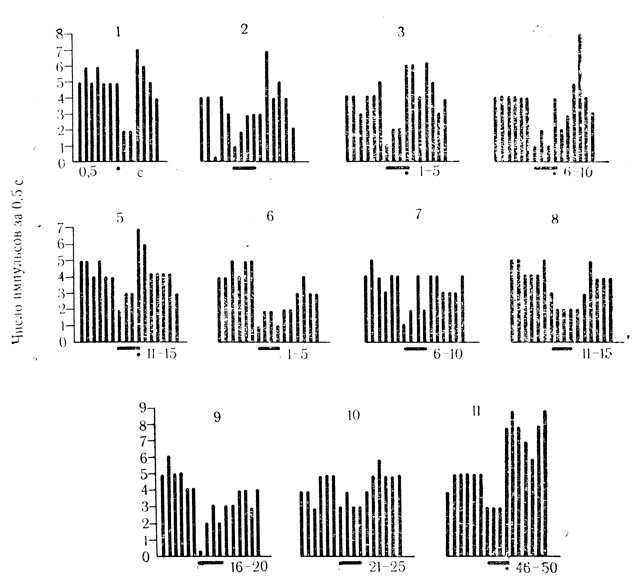
Рис. 20. Растормаживание ответов нейрона слуховой коры на подкрепляющий стимул по мере появления тормозных условных реакций на звук (щелчки - 20 в 1 с) и угашение растормаживающего действия. 1, 2 - контрольные пробы на электрокожное раздражение конечности (точка) и звук (черта); 3-5 - сочетания; 6-10 - пробы на угашение; 11 - восстановление условной реакции повторными сочетаниями. Каждая гистограмма представляет 5 усредненных сочетаний или проб на угашение
Такого рода влияния условной активности нейронов на их безусловные реакции описаны также в структурах ретикулярной формации ствола мозга у крыс (Buresova, Bures, 1965), бледного шара обезьян (Travis e. a., 1968), гиппокампа кролика (Б. И. Котляр, 1969), в зрительной коре кошки (Ю. А. Фадеев, 1970), в сенсомоторной коре кролика (Р. С. Мнухина, 1970) и в моторной коре обезьян (Fetz, Finocchio, 1972) при разнообразных формах выработки условных рефлексов.
Общие принципы структурно-функциональной организации тормозных и растормаживающих путей, известные в настоящее время (Eccles, 1969), позволяют отнести такие нейроны к элементам, использующим аксонные коллатерали возвратного действия через системы тормозных и возбуждающих вставочных клеток и наделенным механизмами растормаживания. Без участия последних трудно объяснить повышение частоты разрядов на подкрепление, которое предваряется угнетением реакции на сигнальный стимул у нейронов с тормозной формой условной активности. Эти наблюдения делают весьма вероятным предположение Eccles (1969) о распространенности механизмов растормаживания в центральной нервной системе.
Торможение ответов на подкрепление у нейронов с активирующими условными реакциями реализуется, по-видимому, механизмами возвратного действия через аксонные коллатерали, терминали которых подходят к тормозным вставочным клеткам, как схематически представлено на рис. 21. Условный разряд "выходного" нейрона (рис. 21, II) активирует тормозные вставочные клетки через возвратные коллатерали и оказывает, таким образом, тормозное действие на этот нейрон в момент прихода по общему пути с "входного" нейрона (рис. 21, I) подкрепляющего компонента реакции. При этом тормозящее влияние условного ответа на подкрепление будет тем лучше выражено, чем интенсивнее проявляется активирующая условная реакция. Е. Н. Соколов (1967) считает такого рода зависимость отличительной чертой механизма возвратного торможения.
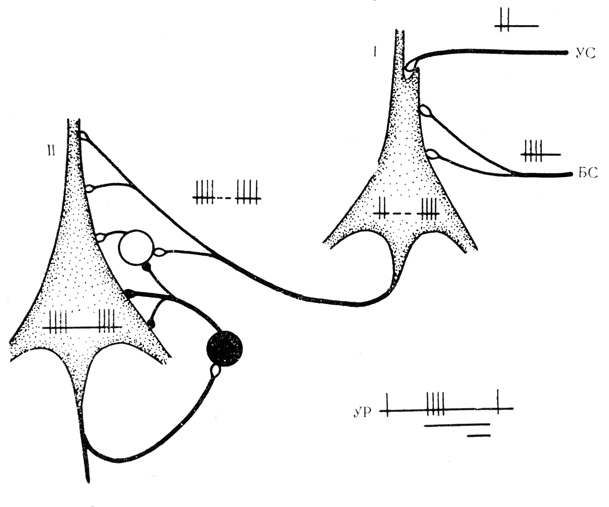
Рис. 21. Схема организации клеточных механизмов торможения подкрепляющего действия. I - 'входной' нейрон, на котором конвергируют условный (УС) и безусловный (БС) стимулы; II - 'выходной' нейрон, па который по общему пути передается сложившаяся связь между сочетаемыми стимулами: УР - условная реакция 'выходного' нейрона. Белый кружок - возбуждающая вставочная клетка, черный - тормозная вставочная клетка. Дополнительные пояснения в тексте
При выработке на "выходном" нейроне тормозной условной реакции сигнал первично интегрированной условной связи приходит сюда с "входного" нейрона через общий тормозной путь, как это представлено на рис. 22. Растормаживание ответов на подкрепление наступает, вероятно, по мере одновременной активации клеток системы растормаживания (терминали аксонов тормозных вставочных клеток подходят к другим тормозным вставочным клеткам) через коллатерали путей безусловного стимула и вставочных клеток возбуждающего действия, также расположенных на входе безусловного стимула (рис. 22, II). При такой организации условный стимул будет вызывать тормозную, а безусловный стимул - активирующую реакцию "выходного" нейрона. Степень растормаживания эффектов подкрепления также обычно зависит от интенсивности предшествующей условной тормозной реакции, но здесь, по-видимому, действуют особенности механизмов посттормозной отдачи. Как показали В. Г. Скребицкий и И. Н. Шаронова (1972), чем более выражены ТПСП в ответ на стимуляцию, тем интенсивнее посттормозная отдача.
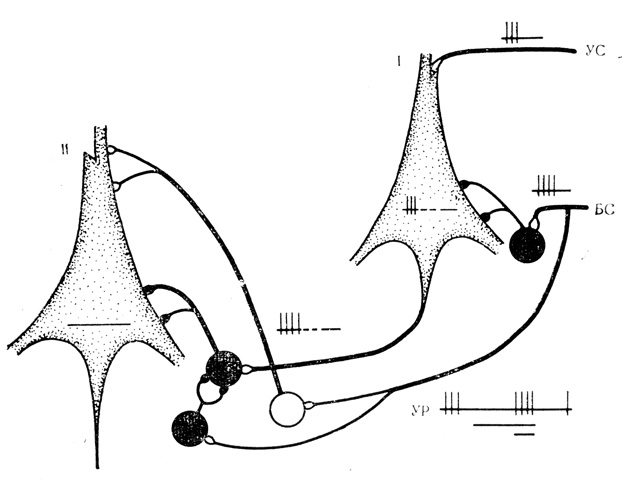
Рис. 22. Схема организации клеточных механизмов растормаживания реакции на подкрепление. I - 'входной' нейрон, на котором конвергируют условный (УС) и безусловный (БС) стимулы; II - 'выходной' нейрон, на который по общему пути передается сложившаяся связь между сочетаемыми стимулами; УР - условная реакция 'выходного' нейрона. Белый кружок - возбуждающая вставочная клетка, черный - тормозная вставочная клетка. Дополнительные пояснения в тексте
Наиболее важен тот факт, что нейрофизиологические механизмы возвратного тормозного действия и растормаживания проявляют условнорефлекторную природу: включаются сочетаниями сигнального стимула с биологически значимым подкреплением и выключаются при его отмене в процессе угашения (см. рис. 19 и 20). По отношению к условному поведенческому акту нейроны, столь эффективно использующие механизмы возвратных тормозных влияний и растормаживания, выступают как функциональные субсистемы с достаточно точной оценкой результатов действия и коррекцией этих результатов, что предусматривается теорией функциональной системы П. К. Анохина (1970).
При выработке поведенческих оборонительно-двигательных актов, как в 1935 г. впервые показал Э. А. Асратян (1953), эффекты торможения подкрепляющего действия сказываются в значительном ослаблении безусловной реакции животного даже на болевые стимулы, если им предшествовали хорошо выраженные условные ответы на сенсорные раздражения.
В заключение следует отметить следующее: наряду с сугубо относительным положением в цепи нейронов клеток с "входными" и "выходными" функциями, как это постулируется нашей исходной гипотезой организации клеточных механизмов условной связи (см. главу I, с. 22), нельзя исключить, что в процессе упрочения целостных поведенческих актов одни нейроны могут преимущественно осуществлять функцию "входных", а другие - "выходных" в пределах каждой структуры мозга, причастной к данному условному рефлексу. В такой ситуации "входные" клетки должны отличаться потенциированными синаптическими входами для сигнального и подкрепляющего стимулов с отчетливым воспроизведением обоих компонентов условной связи. У "выходных" нейронов должны быть потенциированы главным образом входы общего пути для сложившейся ассоциации в целом, возможно, что именно такие клетки наделены механизмами торможения и растормаживания эффектов подкрепления, придающими реакциям фазность, которая способствует эффективной передаче на последующие уровни интеграции, где подкрепляющего действия уже по существу не требуется.
Если такая специализация имеет место в действительности, она складывается, вероятно, только в процессе сочетаний и ограничена набором структур мозга, приспособленных к реализации поведенческих актов вполне определенной биологической значимости.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'