Синаптические механизмы условной связи
Синаптические механизмы образования условных связей на уровне корковых нейронов впервые исследовались в нашей лаборатории Л. Л. Ворониным и Р. Г. Кожедуб с использованием внутриклеточных приемов регистрации (Л. Л. Воронин, 1970в; Л. Л. Воронин, Р. Г. Кожедуб, 1970, 1971а, б). Ранее сходные синаптические реакции зарегистрировали в мотонейронах спинного мозга Spenser с соавторами (1966) и в клетках бульварных ретикулярных ядер Segundo с соавторами (1967) (см. также Segundo, Bell, 1970) в процессе развития привыкания при повторных предъявлениях электрического стимула (клеточные аналоги угашения ориентировочной реакции, или "негативное научение", см. Е. Н. Соколов, 1969) ."'Синаптические механизмы привыкания детально изучены Kandel (1970), Kandel с соавторами (1970) на идентифицированных нейронах брюшного ганглия аплизии, контролирующих ориентировочный рефлекс втягивания жабер.
В модельной ситуации выработки условных связей сочетаниями электрического раздражения двух точек корковой поверхности Л. Л. Воронин и Р. Г. Кожедуб (1971а, б) зарегистрировали (внутриклеточно и квазивнутриклеточно) синаптические реакции нейронов двигательной коры бодрствующих кроликов в зоне моторного представительства передней конечности. Подкрепляющим ("безусловным") стимулом служило пороговое для двигательной реакции конечности раздражение коры через биполярные серебряные электроды, расположенные у места введения регистрирующего микроэлектрода. Эффекты подкрепляющего действия вызывались либо короткой пачкой импульсов длительностью 40-60 мс с частотой 100-200 имп/с, либо более длительной пачкой - до 120-900 мс с частотой 25-50 имп/с. "Условным" стимулом являлись обычно малоэффективные раздражения поверхности коры через такие же макроэлектроды, удаленные от места регистрации каудально на 2,5-12 мм; при этом использовались либо одиночные импульсы длительностью 0,1-0,3 мс, которые предшествовали подкреплению с интервалом 400-800 мс, либо серия из 6-11 таких импульсов и тогда последние 2-3 импульса совпадали с подкреплением. Интервалы между сочетаниями варьировали в пределах от 1 до 120 с.
Подлинность условнорефлекторной природы синаптических реакций клеток идентифицировалась: пробами на неспецифическую сенситизацию ("псевдообусловливание") с раздельным, вне сочетаний, предъявлением "условного" и "безусловного" стимулов; угашением реакции при отмене подкрепления и ее восстановлением повторными сочетаниями; по изменению знака реакции на "условный" сигнал при сочетаниях; по воспроизведению следовой реакции на подкрепление при его пропуске в сочетании.
Результаты исследования показали, что природа и устойчивость изменений постсинаптических реакций клеток по мере сочетаний зависят от степени эффективности подкрепляющего стимула, как это и следовало ожидать, и от длительности интервалов между сочетаниями, что явилось неожиданным и представляет существенный интерес. При небольших межсигнальных интервалах, не превышающих 5 с, и достаточно эффективном подкреплении у большинства нейронов (20 клеток из 28) после нескольких сочетаний появились отчетливые изменения синаптических реакций на сигнальный стимул. Однако эти изменения отличались рядом свойств, присущих неспецифической гетеросинаптической сенситизации клетки: они были неустойчивы и самопроизвольно исчезали спустя примерно 20-25 с; как правило, они не зависели от характера первоначальной реакции той же клетки на подкрепление; сходные изменения возникали и вне сочетаний после нескольких предварительных предъявлений подкрепляющего стимула.
На рис. 24 можно видеть, что контрольное предъявление "условного" стимула вызывает нпзкоаплитудный ВПСП с одним спайковым разрядом, за которым следует длительный и хорошо выраженный ТПСП (А, 1). После нескольких сочетаний с интервалами между ними в пределах 2-4 с амплитуда и длительность ВПСП, вызванного сигнальным раздражителем, существенно возрастают и на его вершине появляется группа спайковых разрядов (А, 2-6). Эти реакции неустойчивы и при тестировании их "условным" раздражением уже на 22-й секунде после сочетания не могли быть обнаружены (Б, 1-5). Сходные по характеру и стабильности изменения постсинаптических реакций могли быть получены при пробах на "псевдообусловливание" (В, 1-6).
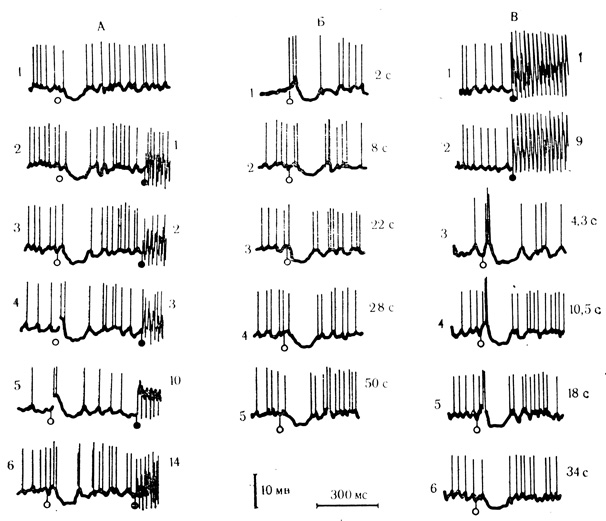
Рис. 24. Синаптические реакции нейронов двигательной коры при коротких (2-4 с) интервалах между сочетаниями (А, Б) и при пробах на 'псевдообусловливание' (В). А: 1 - контрольная реакция на условный стимул; 2-6 - сочетания; показа ю начало длительного (900 мс) безусловного стимула; каждый из импульсов подкрепления вызывает потенциал действия; Б - ответы на условный стимул после серии сочетаний; В: 1, 2 - предъявление подкрепляющего стимула вне сочетаний; 3-б ответы на условный стимул после девяти таких предъявлений. Цифры справа от осциллограмм - порядковый номер сочетаний (А), предъявления подкрепления (В 1, 2) и время (с) после последнего подкрепления (Б, В 3-6). Светлые кружки - отметка условного стимула, темные - подкрепления (Л. Л. Воронин, Р. Г. Кожедуб, 1971 б)
Совершенно иными были изменения постсинаптических реакций клеток при более длительных (12-120 с) интервалах между сочетаниями. В такой экспериментальной ситуации около 43% клеток (9 нейронов из 21) после 10-20 сочетаний проявили синаптические реакции, которые по ряду свойств должны быть отнесены к пластическим сдвигам условнорефлекторной природы. Реакции на "условный" сигнал, как правило, приобретали возбуждающую или тормозную форму в зависимости от знака воздействия на клетку подкрепляющего стимула. При отмене подкрепления синаптические реакции постепенно угасали, но могли быть легко восстановлены повторными сочетаниями, которых теперь требовалось значительно меньше, чем при первых сочетаниях. Повторные серии угашения также требовали меньшего числа предъявлений неподкрепляемого стимула. Наконец, при пропусках подкрепления в сочетаниях в ответ на сигнальный стимул возникала не только условная реакция, но и отчетливая следовая реакция на подкрепление, приуроченная к интервалу отставления "безусловного" стимула.
При возбуждающих формах условной связи в ответ на сигнальный стимул возникали ВПСП, амплитуда которых по мере сочетаний постепенно возрастала до критического уровня генерации спайковых разрядов. При угашении таких условнорефлекторных ВПСП наблюдалось не менее постепенное уменьшение их амплитуды с прекращением генерации потенциалов действия. При этом амплитуды ВПСП обычно снижались до исходного уровня спонтанных колебаний мембранного потенциала клетки ("синаптический шум") без заметного перехода в гиперполяризацию. На рис. 25 видно, что первые предъявления "условного" стимула до сочетаний (Ai3) вызывали небольшое ТПСП, но уже 18-е сочетание способствовало появлению в ответ на "условный" стимул ВПСП небольшой амплитуды и длительности, который предшествовал ТПСП. По мере увеличения числа сочетаний амплитуда ВПСП возрастала, и на ее вершине появился спайковый разряд. При угашении условной реакции амплитуда ВПСП снизилась и достигла уровня "синаптического шума".
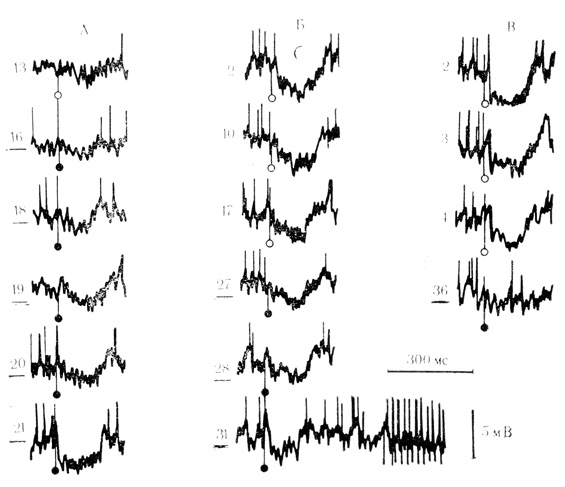
Рис. 25. Возбуждающие условные синаптические реакции нейрона двигательной коры при длительных (20-120 с) интервалах между сочетаниями. Представлены только реакции на условный стимул, за которым следует подкрепление. Начало подкрепляющего стимула видно на Б, 31; каждый импульс подкрепляющего раздражителя вызывает генерацию потенциала действия. Подчеркнутые цифры - порядковый номер сочетания; неподчеркнутые - порядковый номер предъявления стимула при угашении условной реакции. Сочетания - черные кружки, пробы на угашение - светлые. Регистрация через усилитель переменного тока при RC = 0,1 с (Л. Л. Воронин, Р. Г. Кожедуб, 1971 б)
При тормозных формах условной связи сигнальный стимул вызывал отчетливое угнетение механизма генерации потенциалов действия, которое обычно не сопровождалось заметной гиперполяризацией мембраны клетки или достоверным уменьшением ВПСП. При угашении тормозных условных реакций отменой подкрепления спайк-генерирующие способности нейрона полностью восстанавливались. Эти весьма существенные факты еще раз демонстрируют, что по мере выработки условной связи разнообразные нейрофизиологические механизмы, в том числе и механизмы генерации потенциалов действия, способны приобретать новые свойства, обеспечивающие приспособительную реакцию клетки. Примеры выработки и угашения тормозных условных реакций клетки на синаптическом уровне представлены на рис. 26.
Одним из наиболее важных результатов исследования Л. Л. Воронина и Р. Г. Кожедуб (19716) являются данные о том, что угашение условнорефлекторного ВПСП ограничено снижением его амплитуды и не вызывает гиперполяризации клеточной мембраны, т. е. развитие активного тормозного процесса. Подлинная природа этого явления оставалась, однако, неясной, так как на характер изменения полисинаптического ВПСП могло оказать влияние не только повышение тормозящего синаптического притока, но и сохранившийся возбуждающий приток, который компенсирует гиперполяризацию мембраны.
Сходные изменения полисинаптических ВПСП и ТПСП ранее наблюдали Spenser и соавторы (1966) на мотонейронах спинного мозга децеребрированных кошек, а также Segundo с соавторами (1967) на клетках бульбарных ядер ретикулярной формации наркотизированных кошек в процессе выработки клеточных аналогов рефлекса привыкания. Повторные предъявления электрических стимулов вызывали постепенное уменьшение амплитуд ПСП (ВПСП или ТПСП), которые могли быть легко восстановлены предъявлением растормаживающего экстрараздражителя.
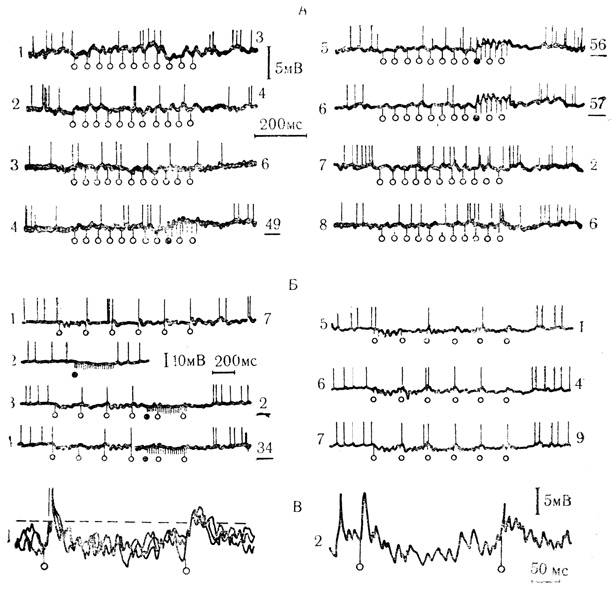
Рис. 26. Выработка и угашение тормозных условных синаптических реакций нейронов двигательной коры. А: 1-3 - ответы на условный стимул до сочетаний: 4-6 - сочетания; 7-8 угашение условной реакции; Б: 1 и 2 - реакции на условный и безусловный стимулы соответственно; 3 и 4 - сочетания; 5-7 - угашение условной реакции. Цифры справа от осциллограмм - порядковый номер сочетаний (подчеркнуты) или предъявления неподкрепляемых условных стимулов. Светлые кружки - условный стимул, темные - начало подкрепления. В: 1 - наложение трех предъявлений условного стимула без подкрепления (ответы на 5-й и 6-й импульсы в пачке, составляющей условный стимул); 2 - ответ в виде частичного потенциала действия на 4-й импульс в пачке условного стимула при угашении. ВПСП, возникающий в ответ на 6-й импульс в пачке (1), превышает пороговый уровень предшествующих ВПСП (1-2), который приводил к генерации потенциалов действия (Л. Л. Воронин, Р. Г. Кожедуб, 1971 б)
Своеобразный характер изменений ВПСП и ТПСП при сдвигах мембранного потенциала клетки, вызванных пропусканием поляризующего тока соответствующего направления через внутриклеточный микроэлектрод и Spenser с соавторами (1966) вычленить деполяризующие и гиперполяризующие компоненты габитуирующих ПСП и на этом основании сделать ряд существенных выводов о природе привыкания. Во-первых, привыкание не связано с появлением нового ПСП и является подлинным уменьшением амплитуды исходного ПСП. Во-вторых, привыкание не связано с изменением знака мембранного потенциала клетки или с изменением порога реакции.
В-третьих, отсутствие достоверных изменений в постсинаптической клетке предполагает пресинаптическую локализацию механизмов привыкания, опосредованных изменениями выброса передатчика из пресинаптических терминалей (см. также Segundo, Bell, 1970).
Весьма важные факты относительно природы угашения и восстановления ВПСП принесли исследования Kandel (1970) и Kandel с сотрудниками (1970), проведенные на идентифицированных нейронах брюшного ганглия аплизии, контролирующих ориентировочную реакцию втягивания жабер. Повторная тактильная стимуляция кожи сифона вызывала угашение рефлекса втягивания жабер, который реализуется расположенными в брюшном ганглии клетками механорецепторов (первичные сенсорные нейроны рефлекса) и жаберными мотонейронами. По мере угашения рефлекса амплитуда ВПСП, регистрируемого в жаберных мотонейронах, постепенно снижалась, такое снижение сопровождалось соответствующим уменьшением числа и частоты спайковых разрядов.
Прямая стимуляция и регистрация одного из механорецепторных нейронов, осуществленная с одновременной регистрацией ответов жаберного мотонейрона, позволили выделить из ряда полисинаптических путей рефлекса моносинаптический путь: каждый спайк первичного сенсорного нейрона вызывал в мотонейроне коротколатентный элементарный ВПСП стабильной формы, который не уменьшался, когда во внешнюю среду вводились вещества, избирательно блокирующие полисинаптические входы. Детальный анализ угашения таких моносинаптических ВПСП повторной стимуляцией позволил заключить, что рефлекс привыкания у моллюсков реализуется без активного торможения постсинаптического мотонейрона и, вероятнее всего, связан с уменьшением выброса передатчика из пресинаптической терминали (Kandel, 1967, 1970).
Одновременная регистрация активности механорецепторных клеток и жаберных мотонейронов представила возможность показать, что восстановление угашенного ВПСП мотонейрона с помощью экстрастимула имело место без изменения частоты импульсации сенсорной клетки. Это наблюдение, по мнению Kandel и соавторов (1970), исключает посттетаническую потенциацию как механизм растормаживающего действия и указывает на то, что восстановление ВПСП растормаживающим стимулом также, по-видимому, опосредовано эффективностью выделенного из пресинаптической терминали передатчика. Kandel и соавторы предполагают, что процессы угашения и восстановления рефлекса втягивания жабер контролируются двумя независимыми механизмами, действующими, однако, в одних и тех же синапсах.
Сопоставление результатов внутриклеточного анализа выработки и угашения условных ВПСП корковых нейронов с механизмами угашения и восстановления полисинаптических и моносинаптических ПСП разнообразных клеточных аналогов рефлекса привыкания указывает на идентичную основу синаптических реакций, выработанных в столь несходных экспериментальных ситуациях и на весьма различных уровнях организации нервных структур. Это обстоятельство позволяет рассматривать элементарные клеточные механизмы выработки условной связи как процесс постепенного и устойчивого повышения эффективности синаптической передачи, основанный, по-видимому, на механизмах синтеза, мобилизации, хранения и выброса передатчика из пресинаптических терминалей.
При этом не следует исключать значение постсинаптических механизмов, таких, например, как изменение чувствительности субсинаптических рецепторов клеточной мембраны к действию передатчика, или обратимые изменения механизма генерации потенциалов действия, которые имеют место при выработке и угашении тормозных условных реакций клетки. Woody и Black-Cleworth (1973) недавно представили экспериментальные данные, полученные на нейронах коронарно-прецентральной коры кошек с предварительно выработанными условными мигательными рефлексами на звук, которые демонстрируют устойчивое повышение сопротивления мембраны клеток, вовлеченных в реализацию условного рефлекса. По мнению этих авторов, локальное и устойчивое повышение сопротивления мембраны и вследствие этого снижение порогов раздражения служат нейрофизиологической основой избирательного действия условного сигнала.
С позиций дальнейших поисков путей и подходов к изучению нейрофизиологических и биохимических механизмов замыкания условных связей мы считаем важным в заключение еще раз обратить внимание на факт неожиданной зависимости пластических свойств клеток от временных интервалов между сочетаниями, которая была обнаружена Л. Л. Ворониным и Р. Г. Кожедуб (19716). Такого рода зависимость трудно объяснить известными сейчас нейрофизиологическими механизмами, и надо, следовательно, допустить, что столь существенные изменения природы синаптических реакций отражают особенности биохимической организации запоминания.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'