Взаимодействие условного и безусловного стимулов на уровне нейрона
Общепринято, что поведенческие реакции на условный стимул формируются по параметрам подкрепляющего рефлекса и воспроизводят эти параметры. Вместе с тем способность нейронов вырабатывать на положительный стимул не только активирующие, но и тормозные условные реакции, указывает на сложные механизмы взаимодействия на клеточном уровне сигнального и подкрепляющего раздражителей и влияние этих факторов на характер условной связи.
Активирующая или тормозная форма условных реакций в корковой зоне проекции подкрепления определяется обычно возбуждающим или угнетающим действием на нейрон безусловного раздражения независимо от знака первоначального ответа на условный стимул. Однако в некоторых случаях и условный стимул, взаимодействуя в процессе сочетаний с подкрепляющим раздражением, влияет на формирование знака условной реакции нейрона. Изредка встречаются клетки, которые не реагировали на подкрепляющий стимул или отвечали на него торможением активности, но по мере выработки условных реакций стали отвечать активацией на оба стимула - условный и безусловный (Е. Л. Полонская, 1967; Г. И. Шульгина, 1967; O'Brien, Fox, 1969b).
Подкрепляющий фактор может оказать влияние ж на другие формы проявления активности нейрона. Так,, по данным В. Б. Швыркова (1968), при сочетании звуковых и световых стимулов с электрокожным раздражением действие подкрепления способствует условнорефлекторной перестройке фоновой активности клеток сомато-сенсорной коры, приближая частоту спонтанных разрядов к структуре ответов на подкрепление. При отмене подкрепления условнорефлекторная перестройка спонтанной активности угасает.
О значении всей функциональной системы условного рефлекса, в которую включаются отдельные нейроны в процессы сочетаний и которая также влияет на характер взаимодействия условного и безусловного стимулов, свидетельствуют наблюдения Г. И. Шульгиной (1968).. Оказалось, что в таком взаимосвязанном ансамбле нейронов каждый из них способен реагировать на одно и то же оборонительное подкрепление в зависимости от модальности условного стимула. У части клеток слуховой и зрительной зоны коры вслед за повышением частоты разрядов в ответ на звуковое условное раздражение наступало торможение активности на подкрепление, тогда как условная активация той же клетки при действии света не препятствовала повышению частоты разрядов и на подкрепление.
Другая часть клеток проявляла реакции обратного"5 знака: при световом условном стимуле ответы на подкрепление тормозились, при звуковом - активировались-В сенсомоторной коре такого типа клетки встречались чрезвычайно редко.
При выработке двигательных условных рефлексов сочетаниями звуковых стимулов с электрокожным раздражением конечности, по нашим данным, активирующая или тормозная форма условных реакций клеток в корковой зоне проекции подкрепления (двигательная кора), как правило, определяется знаком действия на нейрон безусловного стимула. В корковой зоне проекции условного раздражения (слуховая кора) уже только около 2/3 клеток вырабатывают условные реакции по знаку ответа на подкрепление; более 1/3 нейронов проявляют условную активность в соответствии с активирующим или тормозным действием сигнального стимула. У большинства нейронов внутреннего коленчатого тела активирующая или тормозная форма условной реакции формируется под влиянием знака действия на клетку условного сигнала и примерно только 1/4 нейронов вырабатывает условную реакцию в соответствии со знаком действия подкрепления (Е. Л. Полонская, М. Я. Рабинович, 1971, 1972; Rabinovich, Polonskaya, 1973).
Таким образом, наиболее существенные свойства условной связи, ее стабильность, высокая степень афферентной и эфферентной специализации и приспособительная значимость формируются под влиянием воздействия на нейроны подкрепляющего рефлекса. Форма условной реакции - активирующая или тормозная - может определяться как подкрепляющим, так и сигнальным фактором, в зависимости, по-видимому, от соотношения функционирующих на данном нейроне синаптических входов возбуждающего и тормозного действия для каждого из сочетаемых стимулов.
Можно построить схему, удовлетворительно представляющую в общих чертах, каким образом происходит перестройка знака реакции исследуемого нейрона на сигнальный стимул. Если на нейроне число тормозных синаптических входов для подкрепления превышает число возбуждающих входов для сигнального стимула, то при сочетаниях, когда оба стимула конвергируют на общих вставочных клетках тормозного действия (которые в данной ситуации должны преобладать), постепенно будет вырабатываться тормозная условная реакция. В случаях, когда на нейроне число возбуждающих синаптических входов больше для сигнального стимула (как это преимущественно имеет место в зонах его проекции), чем число тормозных входов подкрепления, последующие сочетания и возникающая при этом конвергенция обоих стимулов на общих возбуждающих вставочных клетках приведут к выработке активирующей условной реакции. Более детально эти механизмы схематически изображены на рис. 18.
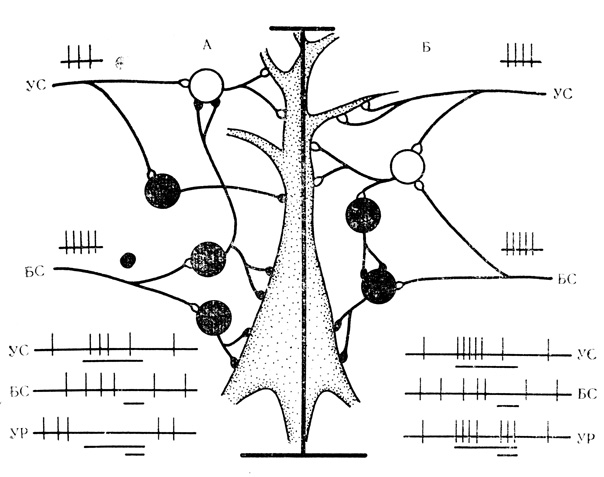
Рис. 18. Схема выработки активирующих и тормозных условных реакций нейрона. Объяснение в тексте
Из рис. 18, А можно составить представление о формировании тормозной условной реакции по знаку подкрепляющего действия. Условный стимул (УС) вне сочетаний активирует нейрон через возбуждающую вставочную клетку (белые кружки); тормозная вставочная клетка (черные кружки на входе УС в данной ситуации неэффективна. Безусловный стимул (БС) вызывает тормозную реакцию нейрона через систему тормозных вставочных клеток. При сочетаниях, когда УС и БС конвергируют на общих вставочных нейронах, БС блокирует возбуждающую вставочную клетку на входе УС, в результате чего УС начинает оказывать на нейрон тормозное действие через ранее неэффективную тормозную вставочную клетку. На рис. 18, Б показано формирование активирующей условной реакции по знаку сигнального стимула. УС вне сочетаний активирует нейрон через прямые афферентные пути и возбуждающую вставочную клетку. БС тормозит активность этого нейрона. При сочетаниях УС через общую возбуждающую вставочную клетку начинает активировать вставочные клетки системы- растормаживания и в результате нейрон активируется как УС, так и БС.
Существенную роль при этом должны играть специальные механизмы растормаживания (система тормозных вставочных клеток, аксонные терминалы которых образуют синапсы на других тормозных вставочных клетках), открытые Wilson, Burgess (1962) в спинальных структурах, и которым Eccles (1969) придает большое значение в деятельности более высоких уровней мозга, включая, по-видимому, неокортекс. Поскольку без привлечения механизмов растормаживания трудно объяснить выработку активирующих условных реакций клеток при тормозном действии подкрепляющего или сигнального стимулов, наши наблюдения могут служить косвенным указанием на универсальность таких механизмов. Об этом свидетельствуют также эффекты растормаживания подкрепляющего действия у нейронов с 5 тормозной условной активностью, о которых будет речь в следующем разделе.
Механизм торможения тормозных вставочных клеток был привлечен В. Г. Скребицким (1967, 1970) и В. Г. Скребицким и И. Н. Шароновой (1972) для объяснения факта подавления тормозных реакций на свет нейронов зрительной коры стимулами незрительных модальностей (звук, электрокожное раздражение конечности, стимуляция ретикулярных структур ствола мозга) и последующего заполнения тормозных пауз импульсными разрядами. Как феномен растормаживания можно рассматривать наблюдения Т. Н. Лосевой с соавторами (1970), показавших, что нейроны, по ряду признаков относимые к вставочным клеткам тормозного действия, сами испытывают торможение в определенных условиях.
Бесспорный интерес представляет тот факт, что в нашем случае механизмы растормаживания имеют условнорефлекторную природу; они формируются только в процессе сочетаний условного и подкрепляющего стимулов, угашаются при отмене подкрепления и невозможны в столь устойчивой форме вне условной связи. Следовательно, можно полагать, что становление и реализация новых, чрезвычайно важных в деятельности мозга механизмов определяется особыми свойствами сочетания в целом и подкрепляющего фактора в частности.
Известно, что для выработки условной связи нейрону необходимы эффективно функционирующие синаптические входы как для сигнального, так и для подкрепляющего стимулов. Эти входы у одной части нейронов функционируют изначально, а у другой части постепенно формируются по мере сочетаний воздействиями подкрепляющего фактора.
Вместе с тем, как это видно из табл. 4, появление у нейронов способности реагировать на оба сочетаемых стимула оказывается недостаточным для образования устойчивой связи между ними. Из числа бимодальных нейронов в двигательной коре выработали условные реакции 78%, в слуховой коре - 59%, а в медиальном коленчатом теле - 50% клеток.

Тадлица 4. Распределение условнореагирующих нейронов в зависимости от характера ответов на контрольные стимулы сочетаемых модальностей
* (Проценты в этой графе посчитаны по отношению к числу нейронов той же строки предыдущей графы.)
Следовательно, в процессе сочетаний нейроны претерпевают какие-то дополнительные пластические перестройки, которым подвержена только часть клеток с бимодальной активностью - заметно большая в зоне проекции подкрепляющего раздражителя и меньшая в зонах проекции сигнального стимула. Таким образом, становление механизмов конвергенции сочетаемых стимулов на одном нейроне следует рассматривать как важнейший предварительный этап в формировании условных связей, обеспечивающих соприкосновение этих раздражителей при первых сочетаниях. В то же время нейроны, которые в контрольных пробах не реагировали на предъявляемые стимулы соответствующих модальностей и были отнесены к числу ареактивных, при последующих сочетаниях оказались способными выработать как активирующие, так и тормозные условные реакции. Таких клеток больше в корковой зоне проекции подкрепления (33% в двигательной коре), чем в зонах проекции сигнального раздражителя (16 и 17% в слуховой коре и внутреннем коленчатом теле соответственно).
Обращает на себя внимание то, что относительное число нейронов, выработавших условные связи (см. табл. 4), возрастает по направлению от внутреннего коленчатого тела (33%) к слуховой коре (44%) и далее к двигательной коре (52%). Это может означать, что активность условнорефлекторной природы распространяется по установленному М. Н. Ливановым (1965) принципу цепных реакций с охватом все большего числа элементов. Такого рода цепной процесс ограничивается, по-видимому, нейронами с тормозной условной активностью, которая вырабатывается на положительный сигнальный стимул одновременно с активирующими условными реакциями (см. рис. 15).
Из анализа данных табл. 4 можно сделать также некоторые выводы о значении дополнительных синаптических входов, которые начинают функционировать по мере сочетаний в зоне проекции подкрепления (двигательная кора) - для сигнального стимула, а в зонах проекции сигнального раздражителя (слуховая кора, внутреннее коленчатое тело) - для подкрепляющего стимула. В группе нейронов, которые в контрольных пробах по ходу выработки условного рефлекса реагировали только на звук, наибольшее относительное число условнореагирующих нейронов оказалось в двигательной коре (56%) и намного меньше - в слуховой коре (40%) и внутреннем коленчатом теле (35%). Наряду с этим в группе клеток, которые в контрольных пробах реагировали только на электрокожное раздражение конечности, наибольший процент условных реакций проявили нейроны слуховой коры (62%) и значительно меньший - нейроны двигательной коры (34%) и нейроны внутреннего коленчатого тела (20%). Надо иметь в виду, однако, что эти соотношения весьма относительны, так как усреднены контрольные пробы на различных этапах выработки и упрочения условного рефлекса, но вместе с тем они указывают на определенные тенденции, которые могут послужить поводом для более достоверного статистического анализа.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'