Динамика условной активности нейронов
Под динамикой условной активности клеток понимают изменения во времени стабильности, интенсивности и характера ответа по мере сочетаний условного и безусловного стимулов и необходимое количество таких сочетаний для появления условной реакции нейрона.
Условные реакции нейронов как активирующего, так и тормозного типа появляются обычно в пределах первых 10-15 сочетаний и относительно быстро (после 30- 50 сочетаний) достигают значимого уровня. Такая сравнительно высокая степень пластичности примерно в равной мере присуща клеткам различных корковых и подкорковых структур мозга при самых разнообразных формах выработки условных рефлексов (Н. Н. Василевский, В. В. Трубачев, 1967; Ф. В. Копытова, М. Я. Рабинович, 1967; В. В. Шульговский, Б. И. Котляр, 1968; Е. Л. Полонская, М. Я. Рабинович, 1971, 1972; Rabinovich, Polonskaya, 1973; Я. Буреш, О. Бурешова, 1965а; Yoshii, Ogura, 1960; Kamikawa e. a., 1964; O'Brien, Fox, 1969a).
Однако приводятся и несколько иные данные. Halas с соавторами (1970) в хроническом эксперименте на кошках с вживленными микроэлектродами вырабатывали классические условные оборонительные рефлексы на звук с электрокожным подкреплением и инструментальные рефлексы избегания электрического стимула на те же звуковые раздражители (тон 1500 кол/с). Регистрировалась активность популяций нейронов (multiple units) в различных образованиях слуховой системы и ретикулярной формации ствола мозга и таламуса.
При выработке простых оборонительных условных рефлексов наиболее рано условные реакции нейронов появляются в ретикулярных структурах ствола мозга, для чего требуется (в среднем для всех животных) 22 сочетания. Немного позже появляются условные ответы в кохлеарном ядре (к 25-му сочетанию) и диффузной таламической системе (срединный центр, центральные латеральное и медиальное ядра таламуса) - к ?30 сочетаниям. В специфических подкорковых образованиях слуховой системы (нижние бугорки, внутреннее коленчатое тело) условные реакции клеток появляются после 48-50 сочетаний, и, наконец, после 70 сочетаний появляются условные ответы в клетках слуховой коры.
Выработка инструментального избегания электрического раздражения требует большего числа сочетаний с уровня кохлеарного ядра, в котором условные ответы клеток в данной экспериментальной ситуации появляются после 38 сочетаний. В нижних бугорках и внутреннем коленчатом теле число необходимых сочетаний возрастает до 56 и 80 сочетаний соответственно. В слуховой коре условные реакции популяции клеток появляются только к 180-му сочетанию. В диффузной таламической системе различия в числе необходимых сочетаний невелики - условные ответы появляются после 36 сочетаний.
В исследованиях Halas с соавторами обращает на себя внимание большая вариабельность (по отношению к отдельным подопытным животным) критических чисел сочетаний, требуемых для появлений условных реакций нейронов как при выработке обычных оборонительных условных рефлексов, так и при инструментальной форме эксперимента. Например, при оборонительном рефлексе условные реакции клеток внутреннего коленчатого тела появляются у разных животных в пределах от И до 76 сочетаний, а слуховой коры - в пределах от 23 до 93 сочетаний. Примерно такие же пределы вариаций критического числа сочетаний имеют место и в других исследованных структурах. По-видимому, усредненные по многим животным количественные значения могут дать только представление о существующей тенденции в последовательности включения отдельных образований мозга в систему условнорефлекторного акта.
Disterhoft и Olds (1972) также исследовали время появления условнорефлекторных изменений ("время обучения") в ряде образований мозга (корковые области, внутреннее коленчатое тело, вентральное, дорсальное, латеральное и заднее ядра таламуса) крыс при выработке пищедобывательных условных рефлексов на звуковые стимулы. Характер условнорефлекторных изменений активности нейронов был сходным- во всех исследованных структурах, но величина и время появления этих изменений существенно различались.
У нейронов внутреннего коленчатого тела и таламической группы ядер первые условные реакции появлялись в пределах 30-40 сочетаний. Корковые нейроны (передняя, средняя и задняя области) начинали реагировать на сигнальный стимул после 60-70 сочетаний.
Появление пищедобывательной реакции требовало несколько больше сочетаний, чем появление условного ответа клеток таламических ядер, но меньше сочетаний, чем это было необходимо для клеток коры.
Такого рода условный поведенческий акт у крыс парадоксально вырабатывался раньше, чем корковые нейроны приобретали способность реагировать на сигнальный стимул.
Вместе с тем авторы отмечают, что максимально выраженные условные реакции клеток во всех исследованных образованиях мозга без различий появляются после 120-140 сочетаний. Однако и при этом величина ответов всегда была относительно большей у клеток таламических ядер, чем в корковых нейроцах. Основываясь на приведенных результатах, авторы полагают, что первичным центром звукового обучения в мозге крысы является таламус и особенно его заднее ядро, через которое, по некоторым анатомическим данным, проходит экстралемнисковый таламо-кортикальный слуховой путь. Следует заметить, что "время обучения" структуры едва ли достаточно для суждения о степени локализации в ней самого обучения. Механизмы включения активности отдельных структур в систему условнорефлекторного акта и последующего выключения из нее весьма динамичны и приурочены к различным этапам становления условного рефлекса (Н. Ю. Беленков, 1969). Об этом свидетельствует как ЭЭГ-анализ условного рефлекса (М. Я. Рабинович, 1967; Т. С. Наумова, 1968), так и данные микроэлектродных исследований, которые будут изложены ниже.
По особенностям динамики параметра стабильности условных реакций в зонах проекции сигнального (слуховая кора и внутреннее коленчатое тело) и подкрепляющего (двигательная кора) стимулов были выделены три типа активности нейронов (М. Я. Рабинович, 1970; Е. Л. Полонская, М. Я. Рабинович, 1971, 1972; Rabinovich, Polonskaya, 1973). К ним относятся нейроны с возрастающей условной активностью, когда условные реакции относительно быстро достигают высокого уровня и клетка стабильно реагирует на сигнальный стимул; нейроны с затухающей условной активностью, отличающиеся постепенным уменьшением ответов на сигнальный стимул до полного прекращения реакций, и в то же время ответы на подкрепление сохраняются неизменными; нейроны с волнообразной условной активностью, которая характеризуется чередованием периодов повышения и снижения стабильности ответов на сигнальный стимул.
В двигательной коре около половины клеток проявило условные реакции волнообразного типа, немного больше 3/4 клеток были с возрастающей условной активностью и около 3/4 нейронов - с затухающими условными реакциями. В слуховой коре было больше клеток с волнообразной активностью (54%), нейронов с возрастающими и затухающими условными реакциями было примерно столько же, сколько и в двигательной коре. Внутреннее коленчатое тело отличается значительно большим числом клеток с возрастающими условными реакциями (40%), около 44% клеток относилось к волнообразному типу реакций и только 16% клеток проявили затухающую условную активность. Примеры трех типов условной активности нейронов представлены на рис. 16.
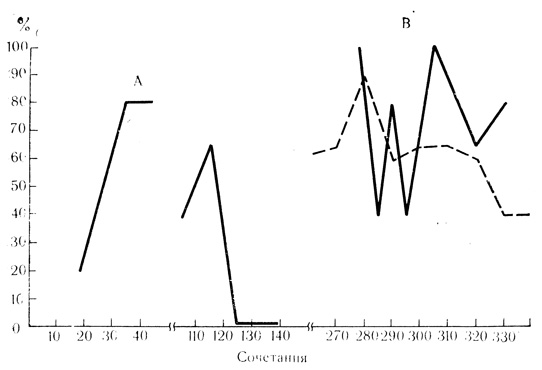
Рис. 16. Динамика условной активности нейронов двигательной коры у бодрствующих кроликов. А - реакции возрастающего типа (клетка на глубине 1230 мкм); Б - реакции затухающего типа (клетка на глубине 590 мкм); В - реакции волнообразного типа (клетка на глубине 800 мкм). Пунктирная линия на В - условные двигательные рефлексы животного. По оси абсцисс - число сочетаний на животном; по оси ординат - процент условных реакций по отношению к 10 сочетаниям (М. Я. Рабинович, Е. Л. Полонская. 1973)
Следует отметить, что возрастающая форма активности наиболее часто встречалась на начальных этапах выработки поведенческого акта.
Вместе с тем при длительной регистрации одного и того же нейрона можно видеть, что один тип условных реакций часто переходит в другой и в ряде случаев уровень ответов клетки на сигнальный стимул снижается на более или менее длительное время. При этом иногда удается проследить весь цикл развития условной активности нейрона, которая относительно быстро достигает значимого уровня положительных ответов на сигнальный стимул и затем убывает (рис. 17). При кратковременной регистрации ответов одного нейрона и сравнительно небольшом числе предъявленных сочетаний обычно улавливается только фрагмент всего цикла развития условной активности клетки, который отражает либо максимальный, либо минимальный уровень цикла. Этим можно объяснить тот факт, что на фоне выработанного у животного условного рефлекса лишь часть клеток реагирует на сигнальный стимул с первого предъявления, а другая часть вырабатывает условные связи как бы заново.
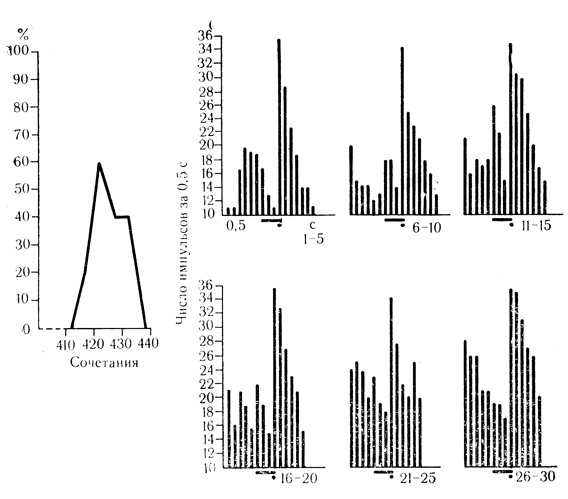
Рис. 17. Затухающая условная реакция нейрона двигательной коры бодрствующего кролика. Клетка на глубине 810 мкм. На графике полный цикл развития условной активности клетки; на гистограммах торможение активирующей условной реакции клетки по мере сочетаний звукового стимула (тон 200 кол/с) с электрокожным раздражением конечности. Каждая гистограмма представляет собой усреднение пяти сочетаний (черта - условный стимул, точка - подкрепление). Гистограмма (здесь и на других рисунках) построены на основании подсчета числа импульсов за каждые 0,5 с в пределах: 2-3 с фоновой активности, 2 с действия сочетаемых стимулов в 2-3 с постстимуляционного фона
Обращают на себя внимание нейроны с затухающей условной активностью. Иногда исчезающие реакции на некоторое время могли быть восстановлены продолжающимися сочетаниями, но вскоре они вновь затухали, проделав один или несколько таких циклов. Обычно из-за неустойчивости условных ответов такие нейроны не привлекают внимания исследователя и относятся к числу невырабатывающих условные связи. Между тем эта своеобразная форма активности клеток отражает, по-видимому, постепенную концентрацию, условнорефлекторных пластических сдвигов в относительно - небольших популяциях нейронов, локализующих выработанный поведенческий акт. Можно допустить, что описанные нейроны имеют отношение к клеточным механизмам хорошо известного феномена концентрации и иррадиации основных нервных процессов, некоторые нейрофизиологические аспекты которого в последнее время рассмотрены А. Б. Коганом (1967).
Интересно, что клетки с затухающей условной активностью не только перестают отвечать на положительный условный стимул, но и отчетливо тормозятся им, по-прежнему реагируя на подкрепление, как это видно на примере нейрона с активирующей условной связью (см. рис. 17). Клетки с тормозной условной связью при затухании активности реагируют на положительный стимул таким же постепенным повышением частоты разрядов. Напомним, что затухающие условные реакции клеток отличаются от известного в лабораториях И. П. Павлова феномена угашения с подкреплением тем, что в данном случае ответы на подкрепление остаются неизменными (см. рис. 17).
Волнообразную форму изменения интенсивности условных реакций клеток двигательной зоны коры отметили O'Brien и Fox (1969a), выделив в процессе сочетаний два периода максимальной активности. Ранний период усиления ответов приходится на 30-60 сочетаний; максимальные реакции позднего периода проявляются в пределах 105-150 сочетаний, после которых наступает некоторое снижение величин ответов, но все же они выше контрольных реакций па тот же стимул до сочетаний.
Тенденция к уменьшению условной активности клеток по мере сочетаний выражена в разных образованиях мозга в неодинаковой степени, что, по-видимому, связано как с особенностями структуры, так и с природой подкрепления, т. е. с биологической значимостью вырабатываемого условного рефлекса. При выработке различного рода оборонительных условных рефлексов отмечено, что нейроны лобной области коры обезьян перестают реагировать па условный и безусловный раздражители сразу же вслед за первыми проявлениями условных разрядов (Jasper e. a., 1960). Нейроны ретикулярной формации ствола мозга кошки и крысы, в которой рано и легко возникают клеточные условные ответы, перестают реагировать на сочетания после 50-60 предъявлений раздражителей (Yoshii, Ogura, 1960; Я. Буреш, О. Бурешова, 1965 а). Тенденцию к уменьшению условной активности клеток ретикулярных структур ствола мозга по мере упрочения условного рефлекса отмечают также Halas с соавторами (1970). Примерно после такого же количества сочетаний уменьшается, а в некоторых случаях прекращается условная активность корковой зоны проекции условного раздражения у кроликов и кошек (В. В. Шульговский, Б. И. Котляр, 1968; Ю. А. Фадеев, 1969). При этом вначале уменьшаются или исчезают реакции нейронов на подкрепление, а затем и на условный стимул, что видно на примерах динамики условной активности ретикулярных клеток, приведенных в работе Yoshii, Ogura (1960).
Наименее выражена тенденция к последующему уменьшению условных реакций в клетках корковых зон проекции оборонительного подкрепления, где условная активность прослежена на протяжении довольно длительного времени (Н. Н. Василевский, 1968; В. Б. Швырков, 1969; O'Brien, Fox, 1969a). Однако и здесь имеют место волнообразно текущие изменения как интенсивности ответа клетки на каждый сигнальный стимул, так и уровня стабильности условных реакций нейрона на протяжении всего периода сочетаний.
Не обнаружено уменьшения условной активности клеток по мере сочетаний в ряде образований мозга (двигательная кора, гиппокамп, гипоталамус, бледный шар, преоптическая область, ретикулярная формация мез- и диэнцефального уровней) при выработке инструментальных условных рефлексов с пищевым подкреплением (Evarts, 1966; Travis, Sparks, 1968; Olds, Hirano, 1969). He проявил себя также феномен уменьшения условной активности в клетках гипоталамуса при использовании в качестве подкрепления стимуляции глюкорецепторов этого образования введением изотонического раствора глюкозы (Б. И. Котляр, Т. М. Ерошенко, 1969). Однако в модельных экспериментах с образованием условных связей сочетаниями звуковых и световых стимулов Morrell (1960) нашел, что нейроны гиппокампа и ретикулярной формации ствола в отличие от клеток зрительной коры перестают реагировать на оба стимула по мере упрочения ассоциации между ними и сохраняют ареактивность на протяжении многих (до 100) последующих сочетаний.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'