Нейроны зрительной коры и наружного коленчатого тела
Исследования принципов конвергенции и взаимодействия стимулов разной афферентной природы на нейронах зрительной коры и наружного коленчатого тела проводились в нашей лаборатории под руководством В. Г. Скребицкого (В. Г. Скребицкий, Л. Л. Воронин, 1965; В. Г. Скребицкий, О. З. Бомштейн, 1967; В. Г. Скребицкий, Л. И. Гапич, 1967; Э. А. Куман, В. Г. Скребицкий, 1968; В. Г. Скребицкий, И. Н. Шаронова, 1972; Skrebitsky, 1969, и др.). В качестве стимулов применялись вспышки света (серии из 5-10 вспышек); тоны разной частоты (200-2000 кол/с); электрокожное раздражение задней конечности, пороговое для двигательной реакции; раздражение ретикулярной формации среднего мозга импульсами длительностью 0,1-0,5 мс с частотой 10 или 100 в 1 с. Все эксперименты проведены на бодрствующих некураризированных кроликах.
Поданным В. Г. Скребицкого (1969), из 236 нейронов зрительной коры 102 клетки (около 40%) реагировали на неадекватный для них звуковой стимул (тон). При этом 33 нейрона (около 32%) из числа отвечавших на звуковые раздражители реагировали повышением частоты разрядов, а 69 нейронов (около 68%) проявляли отчетливое торможение активности при действии звука (табл. 2). Примеры, иллюстрирующие активирующую и тормозную реакции нейронов зрительной коры на звуковые раздражения, представлены на рис. 10, а, б.
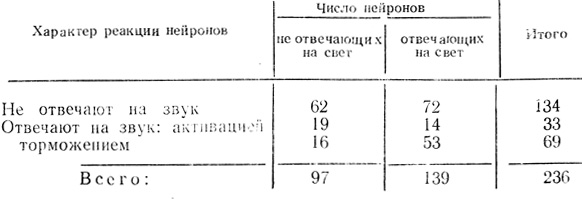
Таблица 2. Распределение нейронов зрительной коры по характеру ответов на световой и звуковой стимулы (по В. Г. Скребицкому, 1969)
Оказалось, что относительное количество нейронов, проявляющих способность к реакциям на неадекватные стимулы, в зрительной коре несколько меньшее, чем в двигательной. Однако основное различие заключается в том, что большинство нейронов двигательной коры реагировало на такие стимулы повышением частоты разрядов, тогда как значительно большая часть корковых клеток зрительной области отвечает на стимулы незрительной модальности торможением активности.
Из табл. 2 следует также, что из 236 нейронов зрительной зоны коры бодрствующего кролика на световые стимулы реагировало 139 клеток. Из этих 139 нейронов 67 отвечали на звук: 14 клеток (21%) активировались звуковыми стимулами и 53 клетки (79%) тормозились. Из 97 нейронов, не реагировавших на свет, 35 клеток отвечали на звуковые раздражители. При этом интересно, что 19 клеток (около 54%) активировались звуком, а 16 клеток (около 46%) отвечали торможением импульсной активности. Эти данные указывают на то, что нейроны зрительной зоны коры, не реагирующие на световые стимулы, отличаются по функциональной характеристике от клеток, одновременно отвечающих и на адекватные, и на неадекватные стимулы. Количество клеток, реагирующих на звуковой стимул, торможением активности, значительно меньше в группе нейронов, не отвечающих на свет, чем в группе нейронов, способных отвечать и на световые, и на звуковые стимулы. По характеру ответов на неадекватные стимулы (преимущественное повышение частоты разрядов) нейроны зрительной коры, не реагирующие на свет, ближе к полисенсорным элементам моторной коры.
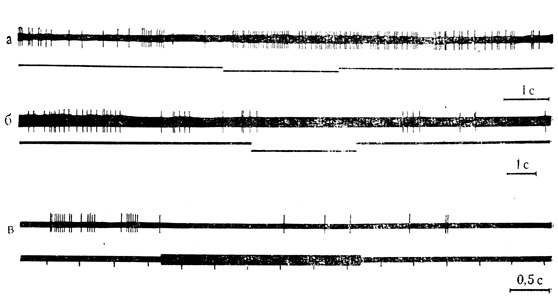
Рис. 10. Ответы нейронов зрительной коры на действие звукового стимула. а - клетка на глубине 0,8 мм; активация нейрона при действии звукового стимула (тон), продолжающаяся в последействии; б - клетка на глубине 1,2 мм, торможение разрядов при действии звукового стимула; в - клетка на глубине 1,16 мм, торможение ответов нейрона на вспышки света при действии звукового стимула, сохраняющееся в последействии. Горизонтальные линии на нижнем луче - отметка звука; вертикальные черточки - отметка света (М. Я. Рабинович и др., 1968)
Более детальный анализ конвергенции и взаимодействия зрительных и незрительных стимулов провели В. Г. Скребицкий и О. З. Бомштейн (1967). Изучалось влияние звуковых сигналов, электрокожного раздражения конечности и раздражения ретикулярных структур среднего мозга на фоновую и вызванную активность нейронов зрительной коры. Из 136 исследованных нейронов значительная часть проявила бимодальные реакции: 41 клетка (30%) отвечала на вспышки света и звуковые стимулы, 37 клеток (27%) - на вспышки и раздражение ретикулярной формации. Из 67 нейронов, испытанных на действие электрокожного раздражения конечности и световых вспышек, бимодальные реакции зарегистрированы у 16 клеток (24%). Из общего числа исследованных нейронов 24 клетки (18%) проявили полимодальные свойства и были способны реагировать ка все испытанные стимулы. Пять нейронов реагировали на один из незрительных стимулов, но не отвечали на вспышки света.
Большинство нейронов (70-75%) реагировало на действие незрительных стимулов торможением активности по примеру ответа на звуковой сигнал, представленного на рис. 10, б. Как правило, одна и та же клетка из числа 24 полимодальных нейронов отвечала на каждый из незрительных раздражителей реакциями одного знака - торможением или активацией. Только у 2 нейронов были зарегистрированы ответы с противоположным знаком.
Механизмы взаимодействия зрительных и незрительных стимулов изучались по изменению характера и структуры ответов на вспышки света при включении одного из разномодальных раздражителей (рис. 10, в, 11). При этом оказалось, что способность к взаимодействию проявили не все нейроны с бимодальными реакциями.- Так, из 41 клетки, отвечавшей на световые вспышки и звук, только 27 клеток (66%) изменяли реакцию на свет при включении звукового стимула. Стимуляция ретикулярной формации оказала влияние на действие вспышек у 26 клеток из 37 (70%), отвечавших на оба стимула. Электрокожное раздражение конечности изменяло реакцию на вспышки света у 88% клеток (14 нейронов из 16, реагировавших на оба стимула). Наиболее эффективным фактором в становлении механизмов взаимодействия выглядит электрокожмное раздражение. Однако такое весьма вероятное заключение требует статистического анализа на значительно большем числе нейронов.
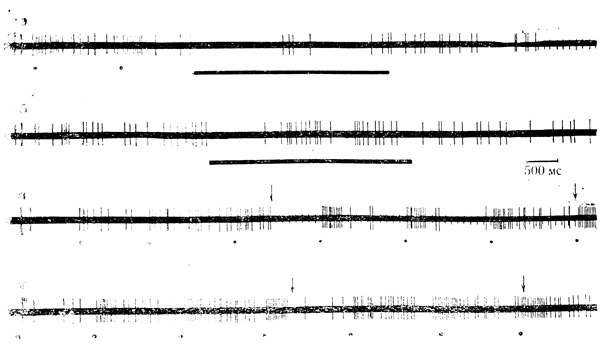
Рис. 11. Влияние незрительных стимулов на фоновую и вызванную (вспышки света) активность нейрона зрительной коры. а - ответы нейрона на вспышки света (обозначены точками), состоят из пачек импульсов и дисперсных разрядов последействия; звуковой стимул (горизонтальные линии) вызывает тормозную реакцию при первом предъявлении; б - угашение тормозной реакции после многократного предъявления того же стимула, торможение выражено при его включении; в - раздражение ретикулярной формации (включение и выключение раздражителя отмечены стрелками) вызывает облегчение реакции на вспышки света, дисперсные v разряды последействия тормозятся; г - электрокожное раздражение конечности (стрелки) несколько уменьшает плотность импульсов в пачке (В. Г. Скребицкий, О. З. Бомштейн, 1967)
Направление изменения реакции клетки на вспышки света при включении стимулов незрительных модальностей обычно соответствовало знаку ответа нейрона на каждый из них. Следовательно, у большинства нейронов реакции на свет действием незрительных стимулов угнетались в пределах тех же примерно 75% клеток (см. рис. 10, в). Ответы на свет у остальных нейронов либо облегчались действием незрительных стимулов (см. рис. 11, в), либо изменения были слабо выражены (см. рис. 11,г).
Торможение фоновых и вызванных вспышками света разрядов клеток зрительной коры действием незрительных стимулов отличается рядом свойств, которые проявляют себя в процессе угашения тормозной реакции повторными предъявлениями тормозящего раздражителя (В. Г. Скребицкий, Л. И. Галич, 1967) и в паттерне синаптических реакций (В. Г. Скребицкий, 1967; В. Г. Скребицкий, И. Н. Шаронова, 1972). Эти особенностш послужили В. Г. Скребицкому поводом выделить описанную форму реакции как "неспецифическое торможение" (Skrebitsky, 1969), которое опосредуется, по-видимому, активностью тормозных вставочных нейронов и механизмами растормаживания (Eccles, 1969), действующими в корковых структурах и о которых еще будет речь впереди (см. главу III, с. 81).
На уровне синаптических реакций (В. Г. Скребицкий, И. Н. Шаронова, 1972) неспецифическое торможение активности нейронов зрительной коры, вызванное действием незрительных стимулов (звук, раздражение ретикулярной формации среднего мозга), выражалось главным образом в угнетении механизма генерации потенциалов действия без заметной гиперполяризации клеточной мембраны (рис. 12, 1). Тормозное действие вспышек света обычно заключалось в появлении коротколатентных (15-30 мс) первичных ТПСП длительностью 20-25 мс, за которыми следовали один или несколько импульсных разрядов и вторая тормозная фаза большей длительности без значительной гиперполяризации мембраны. У части нейронов наблюдались сплошные, относительно длительные ТПСП, которые завершались посттормозной отдачей в виде серии частых разрядов (см. рис. 12, 2).
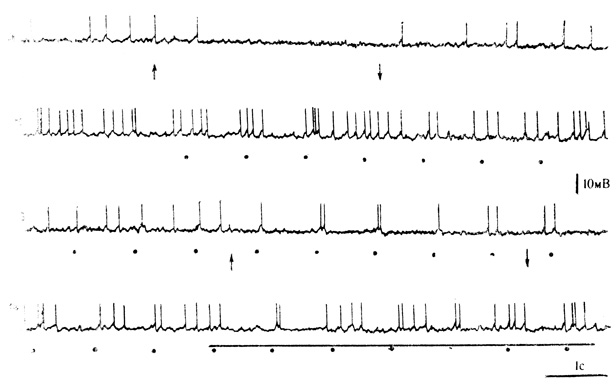
Рис. 12. Синаптические реакции нейрона зрительной коры на зримые и незрительные стимулы (частично внутриклеточная запись). 1 - реакция нейрона на раздражение ретикулярной формации среднего мозга (включение и выключение отмечены стрелками): 2 - вспышки света (точки) вызывают импульсные разряды, за которыми следует длительная тормозная фаза; 3 - раздражение ретикулярной формации (стрелки) вызывает увеличение длительности тормозной фазы реакции на световые вспышки (точки); 4 - звуковой стимул (горизонтальная линия) в начале действия вызывает - угнетение ответов на первые две вспышки (точки) и увеличение длительности тормозной фазы (В. Г. Скребицкий, И. Н. Шаронова, 1972)
Торможение ответов на вспышки света при взаимодействии зрительных и незрительных стимулов проявилось в значительном увеличении длительности второй тормозной фазы и угнетении механизма генерации импульсных разрядов, которые могли иметь место преимущественно при включении незрительного раздражения (см. рис. 12, 3, 4): Облегчающее действие незрительных стимулов сказывалось в уменьшении амплитуды первичных ТПСП и "заполнении" импульсами рой тормозной фазы.
Взаимодействие световых и звуковых стимулов на нейронах наружного коленчатого тела (НКТ) изучали З. А. Куман и В. Г. Скребицкий (1968). Из 100 нейронов - НКТ на вспышки света отвечало 79 клеток. Для большинства клеток ответ состоял из коротколатентного 25 мс) начального разряда с 1-6 импульса, за которым следовали тормозная пауза длительного до 300 мс и поздний разряд (длинно-латентный компонент ответа).
Из 92 нейронов, испытанных на действие звукового стимула, 49 клеток отвечали на звук преимущественно повышением частоты разрядов (37 нейронов активировались звуком, 12 тормозились); 45 нейронов проявили бимодальную активность и отвечали на вспышки света и звуковое раздражение, 34 клетки активировались действием этих стимулов, а у 11 нейронов звуковой раздражитель тормозил активность клетки
Как и у нейронов зрительной коры, только часть клеток с бимодальными реакциями проявила способность к взаимодействию звуковых и световых стимулов. Из 34 нейронов, отвечавших на оба стимула активацией, взаимодействие имело место у 18 клеток (53%); действие звука вызывало облегчение ответов на вспышки света, повышая частоту импульсов начального разряда и длительность позднего компонента ответа, либо "заполнялась" импульсами тормозная пауза (см. рис. 13, А).
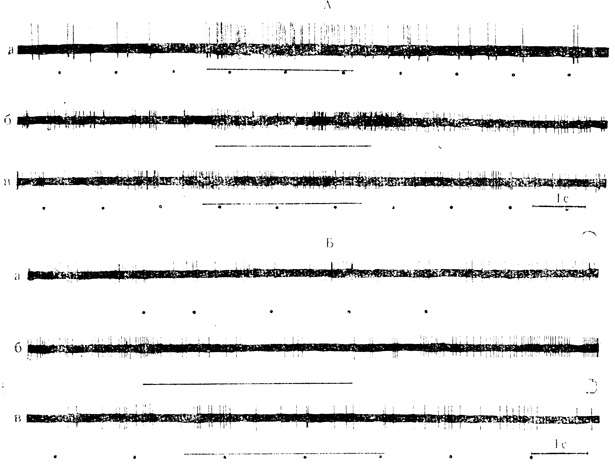
Рис. 13. Влияние звуковых стимулов на активность нейронов наружного коленчатого тела. А - активация звуковыми стимулами: а - нейрон реагирует на вспышки света (отмечены точками) двумя импульсами; действие звука (горизонтальная линия) вызывает значительное облегчение ответов на свет, которое сохраняется в последействии; б - нейрон, реагирующий повышением частоты, разрядов на звуковой стимул: в - действие звука вызывает 'заполнение' импульсами тормозной паузы ответов на вспышки света. Б - торможение разрядов нейрона звуковыми стимулами: а - ответы на вспышки света, состоящие из начального разряда (1-2 импульса), тормозной паузы и относительно длительного позднего разряда; б - торможение разрядов в ответ на действие звука - в - звуковой стимул тормозит главным образом поздний компонент ответов на вспышки света (Э. А. Куман, В. Г. Скребицкий, 1968)
Из 11 клеток, которые на звук реагировали торможением активности, взаимодействие наблюдалось у 10 нейронов и заключалось в торможении реакции на вспышки света: в некоторых случаях уменьшалась частота начального разряда, но чаще всего тормозился поздний компонент ответа (см. рис. 13, Б), что способствовало "выделению" начального компонента.
Таким образом, в процессах взаимодействия световых и звуковых стимулов на нейронах НКТ преобладали факторы облегчающего влияния незрительного стимула на адекватные реакции, вызванные вспышками света, т. е. факторы, противоположные тем, которые действовали в зрительной коре.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'