Нейроны двигательной зоны коры
Первые обстоятельные исследования полисенсорной конвергенции на нейронах двигательной зоны коры принадлежат Buser и Imbert (1964). По данным этих авторов, полученным в острых опытах на кошках, большинство нейронов двигательной зоны коры реагирует импульсными разрядами на электрокожные, зрительные и слуховые раздражения. При этом у животных под хлоралозовым наркозом полисенсорные реакции с коротким латентным периодом проявили 92%, у кураризированных животных - 64% клеток.
При переносе электрода с двигательного поля по направлению к соматосенсорной коре число полисенсорных нейронов значительно уменьшается: у наркотизированных животных до 18%, у кураризированных - до 21%.
У бодрствующих животных число нейронов двигательной зоны коры, на которых конвергируют импульсы различной сенсорной модальности, значительно меньше. А. А. Соколова и Т. Д. Липенецкая (1966) нашли в двигательной зоне коры бодрствующих кроликов только около 36% нейронов (38 клеток из 106), которые были способны реагировать на электрокожное раздражение конечности и звуковые или световые стимулы.
По данным, полученным в нашей лаборатории (Л. Л. Воронин, В. Г. Скребицкий, 1967; М. Я. Рабинович и др., 1968), из 200 нейронов корковой зоны проекции сгибания одной из передних конечностей бодрствующего кролика на неадекватные для них звуковые и световые стимулы реагировали изменением активности 113 клеток (57%) (рис. 3), при этом на свет отвечали 39 клеток (35%), на звук - 25 клеток (22%), на свет и на звук (бимодальные нейроны) реагировали 49 клеток (43%). Одна треть нейронов была способна отвечать импульсными разрядами на стимулы трех модальностей (электрокожное раздражение, свет, звук). Только 9% нейронов (18 клеток) проявляли в ответ на действие зрительных и звуковых стимулов тормозные реакции в виде урежения импульсной активности или ее прекращения.

Рис. 3. Полисенсорные реакции нейронов двигательной коры, а, б - клетка на глубине 2 мм. Ответы на вспышки света с постепенным увеличением активности ('разрабатывание'). Предъявление серии световых вспышек (б) через несколько секунд после предшествующей серии (а) вызывает интенсивный ответ уже на первую вспышку в - клетка зарегистрирована на глубине 1,7 мм. Ответы на звуковой стимул (тон) нейрона с большими амплитудами разрядов. Нейрон с разрядами малой амплитуды на стимул не реагирует. На а и б - отметки вспышки света на нижнем луче. На в - отметки звукового стимула на верхнем луче (М. Я. Рабинович и др., 1968)
Из 70 нейронов двигательной области коры, испытанных на электрокожное раздражение конечности, 38 клеток (54%) проявили повышение импульсной активности. У 18 нейронов (26%) импульсная активность на электрокожное раздражение тормозилась, 14 нейронов (20%) на этот стимул не реагировали. Анализ распределения 125, исследованных нейронов по поперечнику коры (рис. 4) свидетельствует о том, что активность более половины (38 из 65) полисенсорных элементов записана на глубине более 1,6 мм, тогда как среди нейронов, не отвечающих на световые и звуковые стимулы, доля таких глубоко расположенных клеток меньше - 24 из 60 (p<0,05). Максимальное число полисенсорных элементов приходится на уровни 1,6-1,8 мм ниже пиальной поверхности мозга (рис. 4, Б). Эта глубина соответствует примерно нижней части слоя V и верхней части слоя VI по данным Л. А. Кукуева (1940) о ширине клеточных слоев двигательной зоны коры мозга кролика с поправкой на коэффициент сморщивания препарата при его фиксации и обработке. Результаты подсчетов, по-видимому, соответствуют данным Buser и Imbert (1964), обнаружившим полисенсорные клетки в двигательной зоне коры кошки преимущественно на глубине ниже 1 мм от корковой поверхности.
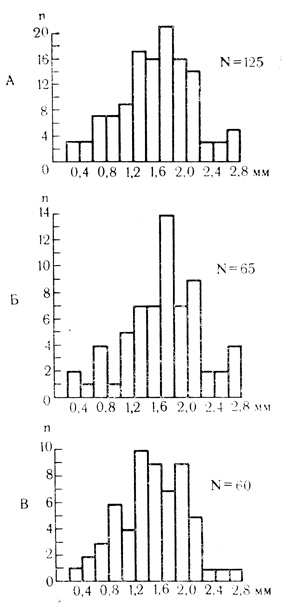
Рис. 4. Распределение полисенсорных нейронов по поперечнику двигательной коры. А - распределение по глубине коры 125 нейронов, подвергнутых действию световых и звуковых стимулов; Б - распределение по глубине 65 нейронов, реагировавших на световые и звуковые стимулы; В - распределение 60 нейронов, не отвечающих ни на один из применявшихся неадекватных стимулов. По оси ординат - число нейронов, по оси абсцисс - глубина регистрации активности нейронов (М. Я. Рабинович и др., 1958)
Интересен характер изменения ответов клеток двигательной зоны коры на длительную повторную стимуляцию неадекватными стимулами (звук, свет). Оказалось, что значительная часть исследованных нейронов проявляет тенденцию к "разрабатыванию", т. е. уменьшению латентных периодов и повышению частоты импульсов в ответ на каждый последующий стимул ритмического раздражителя. Клетки (14), которые вначале не реагировали на данный стимул, начали отвечать на него после повторных применений. Обращает на себя внимание, что постепенное повышение активности, или "разрабатывание", наиболее часто наблюдается у спонтанно малоактивных клеток и обычно сопровождается заметным повышением уровня спонтанной активности нейрона. Из всего исследованного количества нейронов только 3 клетки проявляли быстрое (после 3-4 проб) угашение реакции активации при повторных применениях световых и звуковых стимулов.
Синаптические реакции нейронов двигательной коры у бодрствующих кроликов (внутриклеточное и ("частично внутриклеточное" отведения - см. В. Г. Скребицкий, Л. Л. Воронин, 1966), возникающие в ответ на действие порогового для двигательной реакции электрокожного раздражения конечности, а также звуковых и световых стимулов, проявляли сходные черты с импульсной активностью. Однако количество полисенсорных нейронов могло быть при этом заметно большим, так как часть клеток отвечала (чаще на неадекватные стимулы) подпороговой деполяризацией, не достигавшей уровня генерации пиковых разрядов.
По данным Л. Л. Воронина и В. Л. Эзрохи (1971а), около 80% нейронов (из 70 зарегистрированных) реагировало на электрокожное раздражение передней конечности стабильными коротколатентными (в пределах 7-15 мс) постсинаптическими потенциалами (ПСП). В неповрежденных клетках ответы возникали в виде первичного ВПОП, за которым следовал ТПСП; В ПОП обычно вызывал разряд из одного или нескольких импульсов, а ТПСП эффективно тормозил фоновую активность. Нередко первичный ВПСП был небольшой амплитуды или совершенно отсутствовал и реакция начиналась с коротколатентного ТПСП.
Из 65 нейронов, испытанных на действие звуковых (щелчки) и световых (вспышки) стимулов, 30 клеток реагировали на один из них нестабильными ПСП с длительными (до нескольких сотен миллисекунд) или неопределенными латентными периодами. Часть таких клеток проявила способность к "разрабатыванию" после предъявления одной или двух серий (из 4-10 стимулов каждая) одного и того 'же или разномодальных раздражений; 15 нейронов отвечали на неадекватные раздражители с относительно более стабильными латентными периодами, в пределах 60-150 мс, хотя реакции также возникали не на каждый стимул. Значительный интерес представляют реакции 8 нейронов, которые отвечали на световые или звуковые стимулы стабильными коротколатентными ПОП: 3 клетки - первичными ВПСП, 5 клеток - первичными ТПСП. Латентные периоды реакций на световые вспышки (4 нейрона) были в пределах 20-30 мс, на щелчки (4 нейрона) - в пределах 10-20 мс. Шесть клеток из восьми отвечали также на электрокожное раздражение передней конечности, т. е. проявляли полисенсорные реакции. Как правило, ПСП, возникавшие при действии света или звука, были меньшей амплитуды, чем при реакциях на электрокожное раздражение.
Коротколатентные ПСП свидетельствуют о функционировании у интактных животных весьма коротких путей, способных проводить световые и звуковые сигналы к нейронам моторной коры. К тому же таких клеток оказалось достаточно много (12%) по отношению к общему числу исследованных нейронов. Нейроны моторной коры, проявившие способность реагировать на звуковые или световые стимулы преимущественно нестабильными и длительно латентными ВПСП и ТПСП, описаны ранее Л. Л. Ворониным и В. Г. Скребицким (1967). Записи из этой работы представлены на рис. 5.
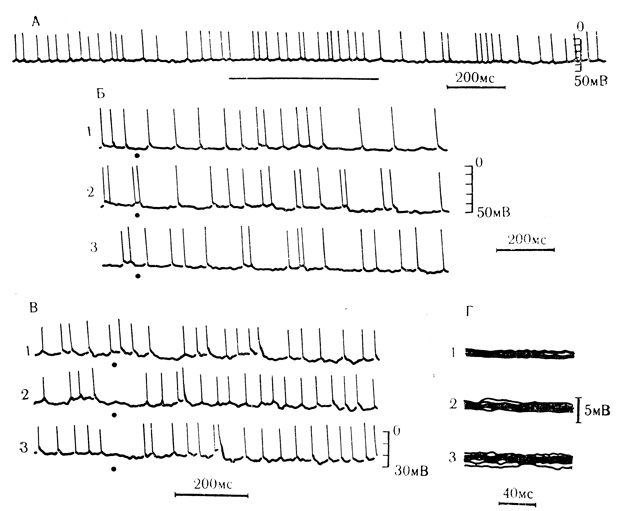
Рис. 5. Синаптические реакции нейронов двигательной коры бодрствующего кролика на звуковые и световые стимулы. А - клетка на глубине 1,3 мм. На включение звукового стимула (горизонтальная черта) отвечает учащением генерации импульсных разрядов, а на выключение - урежением; соответствующие изменения мембранного потенциала слабо выражены; Б - клетка на глубине 1,44 мм. На вспышки света (точки) отвечает усилением синаптической и пиковой активности с длительным латентным периодом (250-300 мс); 1-3 - непрерывная запись; В - клетка на глубине 1,8 мм; в ответ на вспышки света возникают ТПСП; видно уменьшение латентного периода ТПСП при последовательных предъявлениях стимула с частотой 0,9 в 1 с (1-3); Г - клетка на глубине 1,68 мм; возрастание фонового 'синаптического шума' у 'молчащего' (1) нейрона при предъявлении вспышек света (2) и щелчков (3). Запуск развертки синхронизирован со вспышками света и щелчками, идущими с частотой 0,9 в 1 с (Л. Л. Воронин, В. Г. Скребицкий, 1967)
Таким образом, как исследования сотрудников нашей лаборатории, так и многочисленные данные литературы свидетельствуют о том, что значительная часть нейронов моторной коры, в том числе идентифицированные нейроны пирамидного тракта (Д. Н. Ленков, А. А. Пирогов, 1972; Boisacq-Schepens e. а., 1968, Garcia-Rill, Dubrovsky, 1971; Teyler e. a., 1972, и др.), способны отвечать па стимулы различной сенсорной модальности стабильными и коротколатентными реакциями. Более того, для зрительных стимулов у нейронов моторной коры, пирамидных и непирамидных, выявлены рецептивные поля со сложно организованными реакциями на форму раздражителя, его ориентацию и направление движения (Kitsikis e. a., 1969; Garcia-Rill, Dubrovsky, 1971; Teyler e. a., 1972).
Действие поляризующего тока. Не подлежит сомнению, что морфология связей между элементами, образующими данную структуру, так же как и связи с другими образованиями мозга, имеет порвостепенное значение в деятельности полисенсорных нейронов, определяя возможные пути конвергирующей импульсации, характер и латентные периоды их реакций. (Вместе с тем уже то обстоятельство, что количество полисенсорных элементов в одной и той же корковой зоне зависит от
природы веществ, применяемых для обездвиживания животного, как это следует из экспериментов П. Бюзера и М. Эмбера (1964), указывает на значение каких-то функциональных факторов, определяющих в конечном счете способность нейронов к полисенсорным реакциям.
Одним из наиболее изученных функциональных факторов такого рода является поляризация нейронов точно дозированным воздействием на поверхность коры анодом постоянного тока. В лаборатории В. С. Русинова давно и интенсивно исследуется влияние поляризации области проекции передней конечности в моторной зоне коры на формирование двигательных реакций животного в ответ на звуковые и световые стимулы, которые до поляризации таких эффектов не вызывали (поляризационная модель двигательной доминанты). Большой экспериментальный материал лаборатории, обобщенный в монографии В. С. Русинова (1969) и недавнем обзоре (В. С. Русинов, 1972), предваряющем сборник работ лаборатории, имеет прямое отношение к механизмам становления условных связей и будет специально рассмотрен в аспекте анализа эффектов сочетания поляризации нейронов (как модели подкрепления) с действием сенсорных стимулов.
Здесь же важно подчеркнуть возможности, которые представляет анодная поляризация как фактор, способствующий появлению у нейронов полисенсорных свойств. В этой связи интересны результаты, полученные Л. Л. Ворониным (1966) при локальной поляризации отдельных нейронов выходящим током (0,5-5*109 А)
через регистрирующий внеклеточный микроэлектрод. Такая форма опыта позволила упростить экспериментальную ситуацию, устранив возможные эффекты поляризации поверхности коры, неизбежно вовлекающей в процесс относительно большие популяции взаимодействующих нейронов.
Из 70 нейронов двигательной зоны коры бодрствующего кролика, исследованных Л. Л. Ворониным, до поляризации на звуковые и световые стимулы реагировали 36 клеток (51%). На фоне локальной кратковременной (несколько секунд) поляризации каждого элемента число нейронов, реагировавших на эти стимулы, увеличилось до 61 (87%). При этом до поляризации из 36 клеток только на световой стимул отвечало 11 нейронов, на звуковой стимул - 10 клеток, а на световой и звуковой стимулы (бимодальные нейроны) реагировало 15 нейронов. На фоне поляризации через внеклеточный микроэлектрод только на свет реагировало 13 нейронов (небольшое увеличение количества реагирующих клеток), на звуковой стимул отвечало 5 нейронов (уменьшение количества реагирующих нейронов на звук). Зато количество бимодальных нейронов, реагирующих и на звук, и на свет, увеличилось на фоне поляризации почти в 3 раза - с 15 до 43 (рис. 6). Из 70 нейронов 9 (13%) проявили при поляризации тормозные реакции в виде урежения или полного прекращения спонтанной активности. Пластические перестройки активности нейронов оказались довольно сложными.
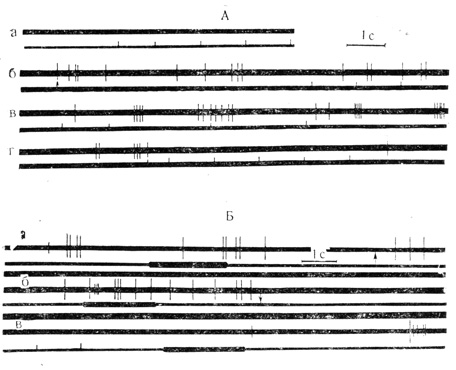
Рис. 6. Ответы на световые и звуковые стимулы нейронов двигательной коры при их локальной поляризации через внеклеточный микроэлектрод. А - клетка на глубине 0,9 мм: а - контрольное предъявление вспышек света до поляризации, реакция на стимул отсутствует; б и в - включение поляризующего тока(5*10 А; отметка стрелкой), вспышки света вызывают разряды клетки; г - поляризующий ток выключен, ответы на вспышки света вновь отсутствуют. Б - на глубине 1,7 мм зарегистрирована активность двух нейронов - с большой и малой амплитудой разрядов: а - контрольное предъявление звукового стимула (тон) ответа не вызывает; б - включение поляризующего тока(5*Ю А; включение и выключение тока отмечены стрелками на а и б); действие звукового стимула вызывает ответ клетки с малой амплитудой разряда; клетка с большой амплитудой также реагирует на звук повышением частоты импульсации; в - после выключения поляризующего тока ответы на звуковой стимул отсутствуют. Отметки стимулов - на нижнем луче (М. Я. Рабинович и др., 1968)
С прекращением поляризации отдельного нейрона обычно исчезали его реакции на неадекватные стимулы в отличие от поляризации поверхности коры, после которой полисенсорные свойства у нейронов сохранялись относительно длительное время (до 20-30 мин), по крайней мере после продолжительного (до 1 ч) действия анодного тока (А. К. Маликова, Р. А. Павлыгина, 1972). Однако Л. Л. Боронину удалось выявить весьма интересный факт: повышение интенсивности поляризующего тока в несколько раз по отношению к оптимальному тормозило реакции нейрона на свет иди звук, предъявляемые на фоне поляризации, но клетки начинали реагировать на эти стимулы в последействии и сохраняли эту способность несколько минут.
Сравнительный анализ эффектов локальной поляризации отдельных нейронов через внеклеточный микроэлектрод с внутриклеточной регистрацией синаптических реакций нейронов моторной коры на фоне анодной поляризации ее поверхности, проведенный Л. Л. Ворониным (1968а), позволяет заключить, что наряду с прямой деполяризацией клеточных тел поверхностная поляризация включает ряд дополнительных факторов, облегчающих ответы на световые и звуковые стимулы. К таким факторам следует отнести вторичные синаптические влияния со стороны клеток во взаимодействующем ансамбле нейронов, одновременно подвергнувшихся деполяризации при приложении анодного тока к поверхности коры (Л. Л. Воронин, 1968а; Purpura, Mc-Murtry, 1965; Gorman, 1967); затем - существенное увеличение эффективности синаптического действия звуковых и световых стимулов, которое связывается с активацией резервных вставочных нейронов; наконец, повышение возбудимости дендритов, которое предположительно имеет место при поляризации корковой поверхности (Л. Л. Воронин, 1968а).
Взаимодействие этих факторов лежит, надо думать, в основе нейрофизиологических механизмов функциональной конвергенции, возникающей в процессе образования поляризационной модели двигательной доминанты. Вместе с тем те же механизмы и, по всей вероятности, ряд дополнительных механизмов могут выступить как звенья, опосредующие какие-то сложные преобразования химических свойств нейрона, характера медиации синаптического действия, что особенно отчетливо выступает при исследовании влияния хлоралозового наркоза на полисенсорную конвергенцию.
Действие хлоралозы. Наиболее результативными оказались исследования синаптических механизмов конвергенции, проведенные в нашей лаборатории под руководством Л. Л. Воронина на нейронах моторной коры наркотизированной хлоралозой кошки (Л. Л. Воронин, Л. И. Таненгольц, 1967, 1969; Л. Л. Воронин, В. Л. Эзрохи, 1972). В качестве стимулов во всех экспериментах применялись: электрокожное раздражение передней контралатеральной и задней ипсилатеральной конечностей (пороговое для вызова реакции сгибания), звуковые щелчки и вспышки света. Одиночные стимулы включались с интервалами не менее 1 в 1 с, сериями из 4-10 предъявлений.
Внутриклеточный анализ показал, что типичные реакции на все стимулы большинства исследованных нейронов (75 клеток из 94) состояли из ВПСП длительностью от 5 до 50 мс, за которыми следовали ТПСП продолжительностью 100-200 мс. Первичные ВПСП приводили к генерации 1-10 пиковых разрядов, а ТПСП эффективно тормозили спонтанную активность и "разряды повреждения" (рис. 7, А, В); 14 нейронов реагировали на стимуляцию первичными ТПСП и, если длительность их не превышала 15-35 мс, за ними следовали стабильные ВПСП. Из общего числа зарегистрированных нейронов 3 клетки отвечали спайковыми разрядами только на электрокожное раздражение передней конечности, одна клетка отвечала лишь на звуковые щелчки и одна клетка - только на световые вспышки. На другие стимулы эти нейроны реагировали подпороговыми деполяризационными колебаниями.
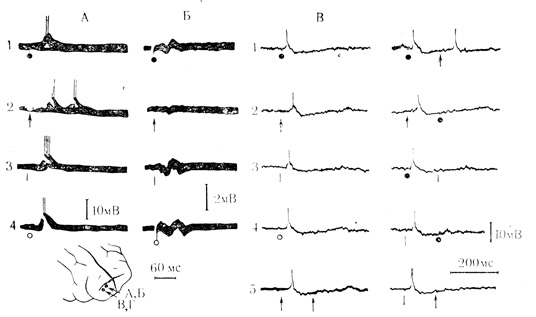
Рис. 7. Постсинаптические реакции двух нейронов (А и В, Г) моторной коры кошки на стимулы различной модальности. Хлоралозовый наркоз. А - ответы клетки на электрокожное раздражение (ЭКР) передней контралатеральной конечности (1, черный кружок); вспышку света (2, стрелка), щелчок (3, черточка), ЭКР задней ипсилатеральной конечности (4, светлый кружок); Б - запись фокальных потенциалов после гибели этой клетки; В - ответы другого нейрона на те же стимулы (1-4). На В5 и Г1 - 5 - предъявление парных стимулов одной или разных модальностей с интервалом 165 мс (см. отметки раздражений). Стрелками на схеме отмечены места регистрации (Л. Л. Воронин, В. Л. Эзрохи, 1972)
Синаптические реакции на все испытанные стимулы относились к коротколатентным, но латентность их варьировала для разных клеток с довольно большим разбросом (табл. 1). Лишь у И нейронов наблюдалась большая стабильность латентных периодов реакций и несколько более высокая вероятность их появления на электрокожное раздражение передней конечности, чем при ответах на световые и звуковые стимулы.

Таблица 1. Латентные периоды постсинаптических реакций нейронов моторной коры наркотизированной (хлоралоза) кошки на стимулы различной модальности (по Л. Л. Воронину и Л. И. Таненголыду, 1969)
Примечание. При вычислении квадратичных отклонений разброс латентных периодов ответа каждого из нейронов на один и тот же стимул не учитывался.
Обращает на себя внимание то, что постсинаптические реакции каждого нейрона на стимулы различной афферентной природы удивительно сходны по форме и временному течению, как это видно на рис. 7, А, В.
Внутриклеточная поляризация токами разного направления (порядка 1-3*10-9А) позволила вычленить "чистые" ВПСП и ТПСП из сложной постсинаптической реакции нейрона и установить, что латентные периоды таких "чистых" ВПСП и ТПСП для каждого из гетеромодальных стимулов совпадали с точностью до 1-3 мс (Л. Л. Воронин, Л. И. Таненгольц, 1969). Это может означать, что возникающие по существу одновременно возбуждающие и тормозные компоненты постсинаптической реакции опосредованы вставочными нейронами возбуждающего и тормозного действия, как это ранее предположили Stefanis и Jasper (1964) на основании анализа антидромных ПСП корковых нейронов пирамидного тракта.
Чтобы выяснить степень влияния хлоралозы на механизмы конвергенции гетеромодальных стимулов, Л. Л. Воронин и В. Л. Эзрохи (19716) исследовали постсинаптические реакции нейронов моторной коры у ненаркотизированных кошек, обездвиженных сукцинил холином. Сравнительный анализ показал, что различия относятся преимущественно к количеству полисенсорных элементов. Так, на электрокожное раздражение передней конечности почти все нейроны (из 83, зарегистрированных в этих экспериментах) отвечали стабильными коротколатентными (менее 20 мс) постсинаптическими реакциями такого же типа, как у животных, находящихся под хлоралозовым наркозом. Около половины нейронов (41 клетка из 83) была способна дополнительно отвечать на вспышки света или звуковые щелчки с короткими и стабильными латентными периодами, до 50 мс, в том числе 6 нейронов реагировали на все предъявляемые стимулы. Хотя форма ПСП была сходной для сигналов адекватных и неадекватных модальностей, однако их амплитуда при действии света и звука всегда была меньшей, чем в реакциях на электрокожное раздражение передней конечности. Подобные различия были отмечены выше у бодрствующих кроликов.
С той же целью сравнительного анализа Л. Л. Воронин и В. Л. Эзрохи (1971а) исследовали конвергенцию стереотипного набора гетеромодальных раздражителей на нейронах сенсомоторной коры наркотизированных кроликов. Оказалось, что у кроликов хлоралоза не вызывает изменений конвергентных свойств в такой степени, как у кошек. Хотя количество полисенсорных элементов, способных отвечать на все испытанные стимулы коротколатентными реакциями, значительно увеличилось по сравнению с бодрствующими животными, однако таких нейронов было всего 15 из 24, зарегистрированных в этой серии опытов, и только у одной трети клеток ответы были более или менее сходными по форме (рис. 8). Остальные нейроны либо не реагировали на все стимулы, либо на один из них отвечали ВПСП, а на другой - ТПСП, либо различались по вероятности появления ответа, который возникал далеко не на каждое предъявление одного из разномодальных стимулов.
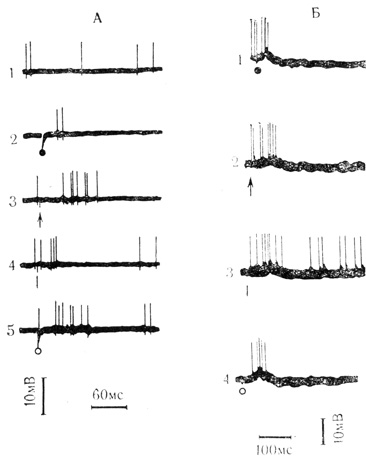
Рис. 8. Реакции двух нейронов (А и Б) сенсомоторной коры кролика на стимулы различной модальности. Хлоралозовый наркоз. А - внеклеточная регистрация. 1 - контрольная запись; 2-5 - ответы на стимулы различной модальности, обозначения модальности раздражителей те же, что на рис. 7; Б - частично внутриклеточная запись: 1-4 - постсинаптические реакции на те же стимулы (Л. Л. Воронин, В. Л, Эзрохи, 1971а)
Взаимодействие стимулов различной модальности у наркотизированных и ненаркотизированных животных. Важные результаты дал анализ взаимодействия стимулов различной сенсорной и биологической модальности на нейронах двигательной коры в условиях хлоралозового наркоза и без него (Л. Л. Воронин, Л. И. Таненгольц, 1967; Л. Л. Воронин, В. Л. Эзрохи, 1971а, б; 1972).
К стимулам биологической модальности мы относим электрокожное раздражение конечностей, так как оно было пороговым и вызывало локальную двигательную реакцию.
У наркотизированных кошек характер взаимодействия не зависел от модальности предъявляемой пары раздражителей, т. е. одномодальные и разномодальные стимулы вызывали однотипные реакции при варьировании последовательности их предъявления. При этом отличительной особенностью выступало так называемое симметричное блокирование, которое заключалось в-том, что примерно в пределах до 200-400 мс после предъявления одного стимула другой раздражитель не способен был вызвать ответ нейрона независимо от модальности как первого, так и второго стимула. Приведенная длительность критического интервала блокирования реакции на второй, тестирующий, стимул наблюдалась у большинства нейронов, но у каждого из них был свой блокирующий интервал с крайними значениями от 100 мс до 1 с.
На рис. 7, Г и рис. 9 представлены эффекты симметричного блокирования при различных интервалах между стимулами. На границах критического интервала постсинаптические ответы на второй стимул были либо ослаблены, либо возникали не на каждое раздражение (см. рис. 9, Г). Только у одного нейрона после предварительного электрокожного раздражения передней конечности реакция на вспышку света, предъявленную с интервалом 165 мс, не блокировалась (см. рис. 7, Г1). Предъявление двух раздражителей одновременно или с интервалами не более 30 мс вызывало ответ, который обычно не отличался от контрольной реакции на каждый из них, или имело место некоторое усиление реакции (см. рис. 9, А3). Следует отметить также, что эффекты блокирования сохранялись, если предъявленному стимулу предшествовал спонтанный постсинаптический потенциал в пределах критического для данной клетки интервала.
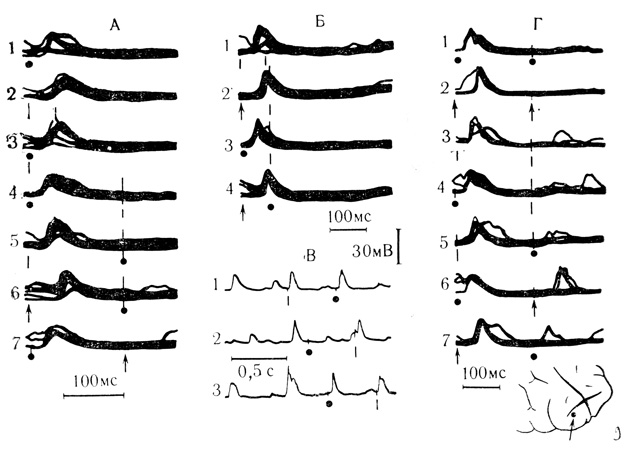
Рис. 9. Взаимодействие стимулов различной модальности на нейроне моторной коры кошки. Хлоралозовый наркоз. А - интервал между стимулами 160 мс: Б - интервал между стимулами 80 мс; В - фрагменты непрерывной записи, интервал между стимулами 430 мс; Г - интервал между стимулами 210 мс. Обозначения модальности каждого стимула те же, что на рис. 7 (Л. Л. Воронин, В. Л. Эзрохи, 1972)
У ненаркотизированных кошек (обездвиженных сукцинилхолином) механизмы взаимодействия разно-модальных стимулов существенно различались. Во-первых, критические интервалы блокирования реакции на второй стимул после предъявления первого были примерно вдвое короче и не превышали 100-200 мс. Часто при таких интервалах реакция на тестирующий стимул была сильнее ответа в его контрольных пробах. Во-вторых, при взаимодействии гетеромодальных раздражителей заметно проявлялось значение модальности каждого из них. Особенно это относилось к электрокожному раздражению конечности, когда оно предъявлялось первым в паре. В таких случаях у большинства исследованных нейронов наблюдалось усиление ответа на последующие предъявления световых или звуковых стимулов или появлялась реакция на эти раздражители, если ее не было в контрольных пробах. Иначе говоря, имело место явление, сходное по механизмам с неспецифической сенситизацией, которая обычно возникает при повторных предъявлениях биологически значимого раздражителя.
При обратном порядке следования стимулов, когда первым предъявлялся звук или свет, ответ на электрокожное раздражение конечности в большинстве проб не изменялся. В-третьих, при одновременном предъявлении разномодальных стимулов или при небольших интервалах между ними общая реакция чаще всего усиливалась.
Интересно, что у кроликов преобладающее значение электрокожного раздражения передней конечности в процессе взаимодействия гетеромодальных стимулов отчетливо выступало даже под хлоралозовым наркозом в отличие от наркотизированных кошек. Однако предшествование электрокожного раздражения в парном предъявлении стимулов здесь вызывало угнетение реакции на последующие звуковые или световые раздражители по типу полного или частичного блокирования. При предъявлении первыми звука или света ответ на электрокожное раздражение конечности не отличался от контрольных проб. При предъявлении двух стимулов одной модальности имело место полное блокирование ответа на второй стимул: исчезала не только импульсная реакция клетки (при внеклеточной регистрации), но и полностью отсутствовал ПСП при внутриклеточных записях. Критический интервал блокирования также не превышал 100 мс.
Эта серия исследований позволила выявить несомненное преобладание стимула биологической модальности (электрокожное раздражение конечности, пороговое для вызова двигательной реакции) над стимулами сенсорных модальностей (свет, звук) при их взаимодействии на нейронах моторной коры, в том числе на идентифицированных клетках пирамидного тракта (Л. Л. Воронин, В. Л. Эзрохи, 19716). Функциональное превосходство электрокожного раздражения конечности наиболее отчетливо проявилось в эффектах асимметричного взаимодействия: реакции на звуковые или световые стимулы усиливались (или угнетались) после электрокожного раздражения конечности, но сами были неспособны оказать влияние на ответ, вызванный электрокожным раздражением, когда они ему предшествовали.
Важно отметить, что элементы преобладания биологически значимого раздражителя носили в себе и симметричное взаимодействие гетеромодальных стимулов. у наркотизированных хлоралозой кошек. Они были выявлены Л. Л. Ворониным (19686) при вычислении частных коэффициентов корреляции латентных периодов реакций на эти раздражители. Оказалось, что достоверная частная корреляция имеет место только для ответов на электрокожное раздражение конечности и какого-либо из сенсорных стимулов (свет или звук). Для электрокожного раздражения и световых стимулов коэффициент частной корреляции обозначался как r = 0,636; для электрокожного раздражения и звуковых стимулов r = 0,332 (P<0,05). Достоверная частная корреляция между латентными периодами ответов на световой и звуковой стимулы отсутствовала (r = 0,098).
Результаты статистического анализа позволили Л. Л. Воронину заключить (Л. Л. Воронин, 1968; Л. Л. Воронин и др., 1971), что электрокожное раздражение конечности не равнозначно звуковому и световому стимулам, как предполагалось ранее (Л. Л. Воронин, Л. И. Таненгольц, 1967). Тем самым предположение Thompson с соавторами (1963) о наличии для разномодальных сигналов общего конечного пути к исследуемому нейрону существенно уточняется: общий путь может иметь место для каждой пары стимулов, которая включает биологически значимый раздражитель. При этом допускается также, что организация такого рода общего пути и его потенциация в определенных условиях (например, хлоралозовый наркоз) могут совершаться в пределах коры механизмами внутрикорковых связей через системы вставочных нейронов возбуждающего действия (Л. Л. Воронин, В. Л. Эзрохи, 19716; Л. Л. Воронин и др., 1971).
Эти результаты хорошо укладываются в предложенную гипотезу организации клеточных механизмов замыкания, в соответствии с которой сложившаяся связь между сигнальным и подкрепляющим стимулом на "входных" нейронах передается на "выходные" нейроны по одному общему пути (см. схему на рис. 2). По-видимому, такой общий путь начинает формироваться уже на этапе становления механизмов конвергенции на уровне нейронов условного и безусловного стимулов.
Заслуживает внимания факт двойственного действия хлоралозы на функциональные свойства клеток и путей к ним. Хлоралоза сенситизирует цепи нейронов, облегчая конвергенцию стимулов различных модальностей на одном конечном нейроне и появление коротко-латентных ответов на эти стимулы. В то же время хлоралоза при взаимодействии гетеромодальных стимулов примерно в 2 раза увеличивает критические интервалы, в пределах которых первый из взаимодействующих стимулов блокирует второй в общем пути. "Хлоралозовый парадокс" отражает, вероятно, различную химическую (медиаторную) организацию синаптических входов для изолированно действующих стимулов и для общего пути, когда эти стимулы взаимодействуют.
Недостаточно выясненными остаются пути проведения звуковых и световых стимулов к нейронам моторной коры, способным реагировать на них стабильными и коротколатентными ответами. Приводятся экспериментальные доказательства в пользу проведения гетеромодальных стимулов через интраламинарные ядра и срединный центр таламуса (Albe-Fessard, 1967). Для зрительных стимулов показана возможность проведения к моторной коре через верхние бугры четверохолмия (Ascher, Gachelin, 1963; Dubrovsky, Garcia-Rill, 1971). Нейроны, реагирующие с относительно короткими латентными периодами на свет и звук, обнаружены в ядре Бурдаха (Т. С. Наумова, Н. А. Ласточкина, 1966).
Имеются морфологические доказательства прямых связей между нейронами внутреннего коленчатого тела, передних и задних бугров четверохолмия и клетками моторной коры (С. Б. Дзугаева и др., 1969). Показаны также прямые связи, идущие от сетчатки через наружное коленчатое тело и лучистый венец к нейронам двигательной коры (В. Г. Скребицкий, Е. Г. Школьник-Яррос, 1964).
Недавно В. С. Русинов (1972) сообщил о прямых связях элементов ретикулярной формации мозгового ствола с нейронами моторной коры, обнаруженных его сотрудниками М. А. Рябининой и Э. Н. Панаховой. Раздражение ретикулярных структур одиночными импульсами, пороговыми для вызова движения передней конечности, вызывали у некоторых нейронов моторной коры (их менее 1%) активирующие и тормозные реакции с латентными периодами 4-5 мс. Эти связи В. С. Русинов относит к моносинаптическим, учитывая время проведения разряда и синаптическую задержку.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'