Глава VI. Взаимодействие специфической и неспецифической афферентации при формировании дифференцировочного торможения
В школе И. П. Павлова было сделано предположение о том, что дифференцировочное и угасательное торможение формируется с участием как корковых, так и субкортикальных структур мозга. И. П. Павлов полагал, что дифференцировочное торможение "развивается в соответственном анализаторе, где происходит анализ раздражителей" (И. П. Павлов, 1949, т. III, стр. 149). В то же время И. П. Павлов отмечал, что торможение не локализуется лишь в анализаторе, воспринимающем дифференцировочный сигнал, а распространяется на рабочие аппараты эффекторного отдела дуги условного рефлекса. Он писал (1932): "Надо представить, что с пунктов искусственных условных раздражителей развивается сильное торможение на весь подкорковый пищевой центр с его обоими главными компонентами - секреторным и двигательным, а также и на соответствующий отдел коркового двигательного анализатора" (И. П. Павлов, 1949, т. III, стр. 419).
В дальнейшем ученики и последователи И. П. Павлова развивали разные стороны этого цельного представления о торможении как процессе, охватывающем систему структур мозга. П. С. Купалов и А. М. Ушакова (1931), А. А. Травина (1954), П. С. Купалов (1960) считали, что внутреннее торможение может возникать как в одном, так и в двух корковых концах взаимодействующих анализаторов. Б. П. Бабкин (1904), Г. П. Зеленый (1907), Л. Г. Воронин (1952), А. И. Ройтбак (1960), У. Г. Гасанов (1962) и другие придерживаются точки зрения, что торможение локализуется лишь в анализаторе, к которому адресуется дифференцируемое раздражение. Э. А. Асратян (1955, 1959) полагает, что торможение формируется где-то на путях между корковым концом условного анализатора и корковым представительством пищевого центра. Группа исследователей (И. Я. Перельцвейг, 1907; Н. А. Кашерининова, 1908; П. К. Анохин, 1932, 1958а; Н. А. Рокотова, 1953; И. И. Короткий и М. М. Суслова, 1955; Б. И. Ходоров, 1955; Г. В. Скипин, 1956; Р. Г. Зевальд, 1958; М. А. Панкратов, 1960) отрицает представление об анализаторной локализации торможения и отстаивает мысль о локализации торможения в эфферентной части дуги условного рефлекса. Вместе с тем, П. К. Анохин (1958а) полагает, что формирование дифференцировочного или угасательного торможения на базе пищевого рефлекса протекает по принципу одновременного создания биологически отрицательной реакции "трудного конфликтного состояния". Формирование внутреннего торможения на базе оборонительной реакции П. К. Анохин рассматривает как прямо противоположное явление: "Устранение выработанной отрицательной реакции животного или его "трудного состояния" через возбуждение положительных реакций не имеет конфликтного характера и потому маловероятно, чтобы оно осуществлялось с помощью одних и тех же механизмов, а именно с помощью активного внутреннего торможения" (П. К. Анохин, 1958а, стр. 156). И далее: "Переход от биологически отрицательной реакции к биологически положительной происходит без активного торможения в том виде, как мы его знаем на примере общеизвестного внутреннего торможения, и, следовательно, конкретный физиологический механизм этого перехода должен стать очередной задачей физиологических исследований" (там же, стр. 158).
По-существу, все электрофизиологические эксперименты подтвердили суждение И. П. Павлова, высказанное им на одной из сред, что дифференцировочное торможение есть смешанное безусловно-условное. Однако механизм и локализация тормозного процесса остаются до настоящего времени предметом разногласий. До настоящего времени нет еще единого мнения даже о том, какое электрографическое выражение имеет дифференцировочное торможение, формирующееся на базе разных условных рефлексов, зависит ли это выражение и в какой степени от характеристики сигнальной стимуляции. В связи с этим нет и общепризнанных суждений о том, как оценить ту или иную электрическую реакцию, которая сопровождает дифференцировочное торможение. Следуя представлениям о том, что медленные волны отражают собой тормозное состояние мозгового субстрата (Jasper, 1941; Gibbs, Gibbs, 1950, 1952, и др.), исследователи искали и находили медленные колебания потенциала в момент осуществления дифференцировочного торможения, вырабатываемого на базе и пищевых (Н. Н. Любимов, 1960) и двигательных (Jus, Jus, 1956, 1959а, b; М. Я. Рабинович, 1958; Roy и сотр., 1961, и др.) рефлексов, однако эти данные не нашли полного подтверждения в самые последние годы. В эти годы появились наблюдения, свидетельствующие о том, что в момент действия отрицательного, дифференцировочного стимула, вопреки ожидаемым медленным волнам, появляется, как правило эффект десинхронизации, сходный с тем, который сопровождает действие условного оборонительного сигнала (Ю. Г. Кратин, 1960; Р. С. Мнухина, 1963, и др.). Как понять наблюдаемое явление? Очевидно, ответ на этот вопрос следует искать в тщательном изучении электрических процессов на разных уровнях анализаторов, а также в тех структурах, которые к ним не причисляются. Нам думается, что этому поможет и изучение динамики параметров вызванных ответов, сопровождающих формирование дифференцировочного торможения. Такое исследование необходимо вести на базе разных условных рефлексов, с использованием разных сигнальных раздражений и на разных животных.
Наше исследование в этой области ограничилось изучением динамики электрических процессов у собак при формировании дифференцировочного торможения, вырабатываемого на непрерывные и ритмические звуковые раздражения разной интенсивности.
Изучение электрической активности мозговых структур при формировании дифференцировочного торможения показало значительные различия в сдвигах ЭЭГ на разных этапах образования дифференцировки. Кроме того, электрографическое выражение этого вида внутреннего торможения в значительной мере зависело от интенсивности стимула, на который происходила выработка тормозной реакции.
Начнем описание с изменений ЭГ, сопровождающих формирование дифференцировочного торможения на подпороговые и пороговые (в смысле вызова электрографической реакции) звуковые раздражения. На рис. 47-48 представлены записи ЭГ корковых и бульбарных структур собаки № 4, сделанные в процессе выработки дифференцировочного торможения на тон 100 гц 59 дб. Видно, что действие этого сигнала в начале выработки на него дифференцировочного торможения (см. рис. 47, а) сопровождается двигательной реакцией и вызывает генерализованное снижение электрической активности в структурах продолговатого мозга и эффект десинхронизации на уровне большинства корковых зон. В сенсомоторной области коры регистрируется резкая экзальтация (в 2-3 раза) электрических колебаний, которая в конце действия отрицательного сигнала еще больше усиливается; при этом усиливаются и осцилляции в кохлеарных ядрах и коре эктосильвиевой извилины. На следующем этапе начинают регистрироваться изменения ЭГ, приуроченные к системе слухового и кожно-двигательного анализаторов. На рис. 47, б представлено 28-е применение дифференцировочного стимула, которое сопровождается двигательной реакцией и вызывает изменения электрических потенциалов, сходные с наблюдаемыми при осуществлении условного рефлекса в первый период концентрации, а именно избирательный рост электрической активности во взаимодействующих анализаторах, отчетливо выраженный в первичных переключательных реле слухового анализатора (кохлеарных ядрах) и корковом конце кожно-двигательного анализатора (сенсомоторной зоне коры). В ядрах нежного и клиновидного канатиков и нижнем центральном ядре ретикулярной формации наступает снижение электрических колебаний. В корковых структурах регистрируется усиление процессов, выражающееся либо в возрастании только частоты колебаний (слуховая область коры), либо в возрастании их ритма и амплитуд (зрительная область коры). По мере дальнейшего неподкрепления этого сигнала сначала исчезает моторная реакция, а затем и ее электрографические признаки. На рис. 48 видно, что лишь в первый момент действия дифференцировочного стимула (43-е применение) наступает некоторое возрастание электрических осцилляций, заметное в сенсомоторных зонах коры и бульбарных структурах мозга, а затем происходит восстановление исходного уровня процессов, и к 2-3 сек действия звукового сигнала активность несколько снижается. В конечном счете, дифференцировочный раздражитель перестает вызывать изменения электрических потенциалов как в структурах коры головного мозга, так и в ядрах ствола.
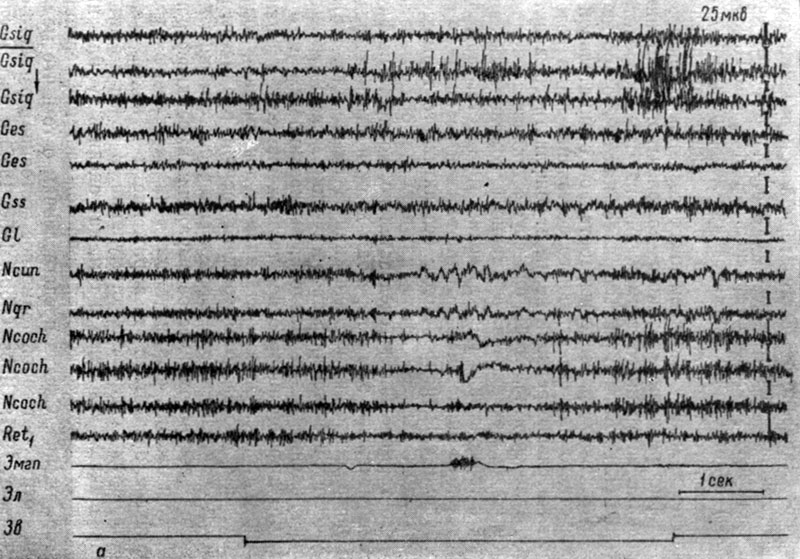
Рис. 47. Изменения ЭГ коры большого мозга и образований продолговатого мозга собаки № 4 в процессе образования дифференцировки на тон 100 гц 59 дб. а - 23-е (опыт № 68 от 27/IV 1959); б - 28-е применение дифференцировочного раздражения (опыт № 73 от 8/V 1959 г. - по Т. С. Наумовой, 1962б)
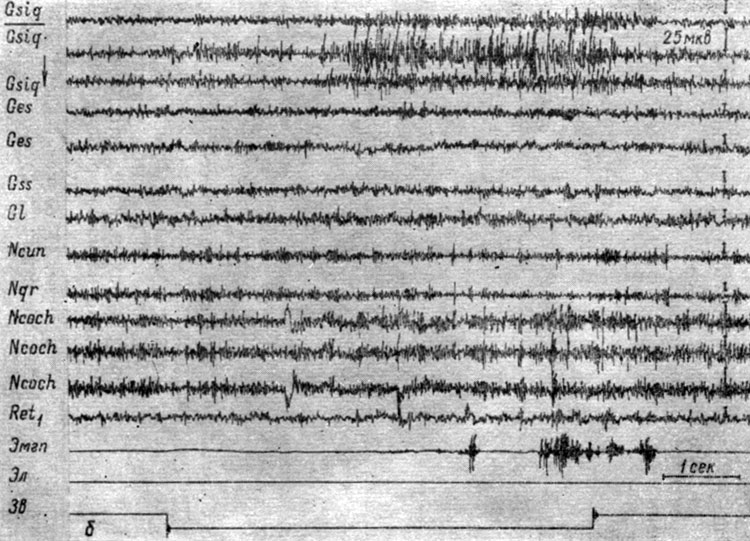
Рис. 47. Изменения ЭГ коры большого мозга и образований продолговатого мозга собаки № 4 в процессе образования дифференцировки на тон 100 гц 59 дб. а - 23-е (опыт № 68 от 27/IV 1959); б - 28-е применение дифференцировочного раздражения (опыт № 73 от 8/V 1959 г. - по Т. С. Наумовой, 1962б)
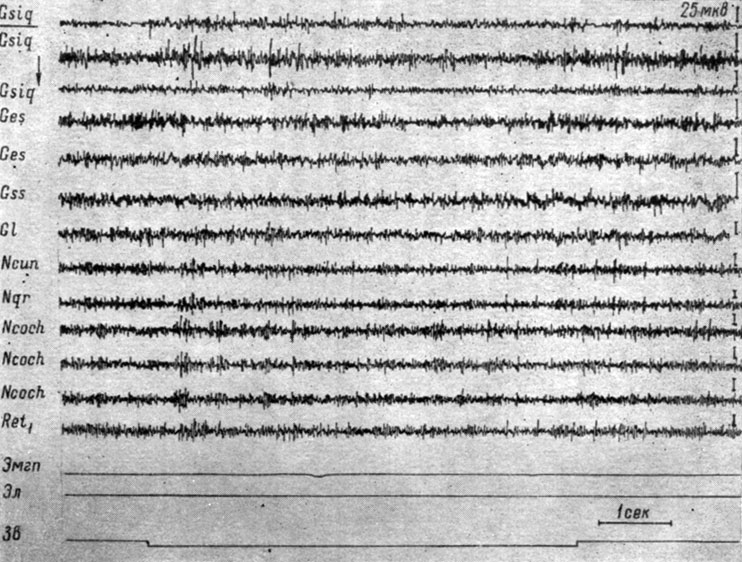
Рис 48. Изменения ЭГ коры большого мозга и образовании продолговатого мозга собаки № 4, наступающие в период упрочения дифференцировки на тон 100 гц 59 дб. (опыт № 87 от 6/VI 1959 г. - по Т. С. Наумовой, 1962б)
Иными словами, формирование дифференцировочного торможения проходит все те стадии, которые сопровождали образование условного оборонительного рефлекса, вплоть до того момента, когда была введена дифференцировка. После этого наступает период, в который действие отрицательного сигнала перестает вызывать видимые изменения ЭЭГ.
Та же последовательность фаз была обнаружена при изучении электрических процессов на других уровнях мозга. У тех животных, у которых I стадия выработки рефлекса отсутствовала, формирование дифференцировочного торможения также не проходило стадию генерализованного снижения электрических колебаний, и двигательная реакция на первых этапах формирования отрицательного рефлекса сопровождалась общим усилением электрических процессов.
Таким образом, тоновые раздражения подпороговой и пороговой интенсивности, электрографическая реакция на которые до выработки дифференцировочного торможения легко угасает, будучи сигналами отрицательной реакции, сначала вызывают движения конечности и сдвиги ЭЭГ, последовательно повторяющие все этапы формирования условных рефлексов, а затем не сопровождаются видимыми изменениями электрической активности. Движения конечности исчезают раньше электрографической картины замыкания.
Если формирование дифференцировочного торможения происходит на звуковые раздражения, по интенсивности значительно превышающие пороговые значения, динамика процессов, сопровождающих эти отрицательные реакции, значительно отличается от только что описанной, но и в этом случае торможение формируется постепенно, проходя те же этапы, которые сопровождают организацию условнорефлекторной реакции на сигналы надпороговой интенсивности.
В начале выработки дифференцировочного торможения на такого рода звуковые раздражения возникают (рис. 49, а - 16-е применение звуковых щелчков в ритме 20 в 1 сек, интенсивность 63 дб у собаки № 4) изменения электрической активности, подобные наблюдаемым при осуществлении условного оборонительного рефлекса на аналогичные звуковые сигналы в стадии концентрации, а именно вслед за генерализованным снижением электрических осцилляций, наиболее отчетливо выступающим в структурах ствола (до 6-12 мкв), регистрируется значительный рост частот и амплитуд электрических колебаний в кохлеарных ядрах (до 20 мкв) и сенсомоторных зонах коры (до 50-75 мкв в записях накладными электродами). В ядрах нежного и клиновидного канатиков, нижнем центральном ядре ретикулярной формации имеет место снижение электрических потенциалов. В большинстве корковых зон незначительно выражен эффект десинхронизации. Следование ритму раздражения нечеткое. В начале выработки дифференцировки эти изменения сопровождаются двигательной реакцией, затем последняя исчезает.
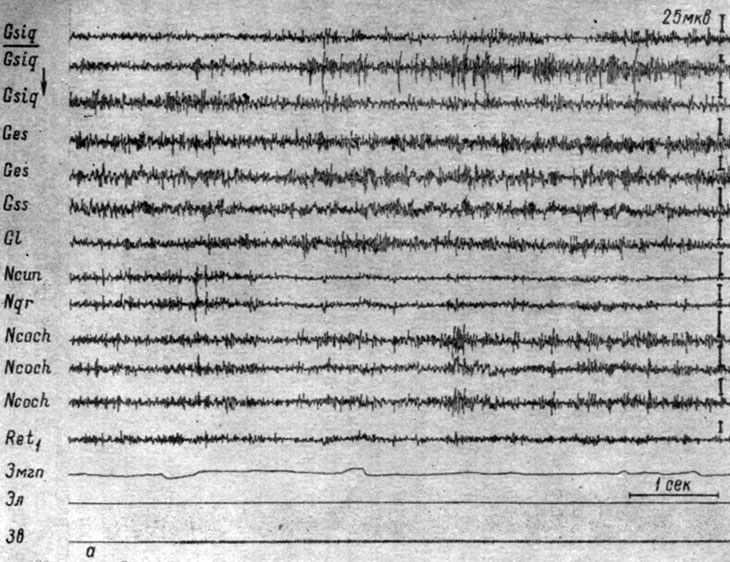
Рис. 49. Изменения ЭГ коры большого мозга и образований продолговатого мозга собаки № 4, наступающие в процессе формирования дифференцировки на щелчки (20 в 1 сек, 63 дб). а - 16-е (опыт № 93 от 2/VII 1959 г.); б - 27-е применение дифференцировочного раздражения (опыт № 104) от 25/IX 1959 г. - по Т. С. Наумовой, 1962б)
На приведенном рисунке в ответ на действие дифференцировочного стимула локальная реакция конечности на звук полностью отсутствовала, но общее движение животного еще имело место (смещение линии ЭМГ). По мере выработки и упрочения торможения отсутствие оборонительной реакции на дифференцировочный сигнал начинает сопровождаться значительным и генерализованным по структурам ствола мозга снижением электрической активности (до нескольких микровольт), длящимся и после окончания раздражения (см. рис. 49, б). Снижение активности наблюдается также в сенсомоторной (до 10-12 мкв) и зрительной (до 8-10 мкв) областях коры. В других корковых зонах (эктосильвиевой и супрасильвиевой извилинах) происходит некоторое нарастание уровня возбуждения, выражающееся в незначительном росте частоты колебаний и нечетком следовании ритму раздражений. Детальный анализ динамики параметров вызванных ответов приводится в конце этого раздела.
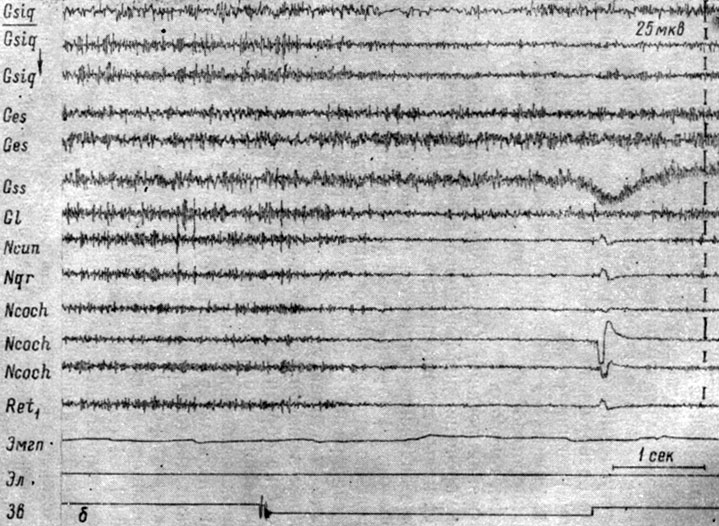
Рис. 49. Изменения ЭГ коры большого мозга и образований продолговатого мозга собаки № 4, наступающие в процессе формирования дифференцировки на щелчки (20 в 1 сек, 63 дб). а - 16-е (опыт № 93 от 2/VII 1959 г.); б - 27-е применение дифференцировочного раздражения (опыт № 104) от 25/IX 1959 г. - по Т. С. Наумовой, 1962б)
Таким образом, более сильные (в смысле вызова электрографической реакции) звуковые стимулы, на которые до сочетаний не удается в полной мере угасить электрографическую реакцию, в качестве дифференцировочных сигналов вызывают генерализованное снижение электрической активности в структурах ствола мозга и реакцию десинхронизации в коре (при записи ее ЭЭГ накладными электродами) и в случае ритмических раздражений следование ритму стимуляции.
Описанные особенности изменений ЭГ характерны и для структур других уровней нервной системы.
Так как генерализованное снижение электрической активности наступает и в ответ на действие физиологически сильных звуков до превращения их в сигналы положительной или отрицательной реакции, можно думать, что такое же электрографическое выражение дифференцировочного торможения на стимулы надпороговой интенсивности зависит от действия раздражителя как такового.
Диффузная реакция (депрессия электрической активности) по мере упрочения дифференцировки, вырабатываемой на звуковые сигналы, по интенсивности значительно превышающие пороговые значения, становится все менее выраженной, и, таким образом, электрографические картины, сопровождающие осуществление дифференцировочного торможения на звуковые раздражения разной интенсивности, становятся сходными. Это положение наступает у разных животных при разном количестве применений тормозного стимула.
Интересно обратить внимание на то обстоятельство, что по мере применения тормозного раздражителя уменьшение уровня диффузных изменений электрической активности выражается не только в характере сдвигов суммарной ЭЭГ, но также в исчезновении аналогов вызванных ответов из "незаинтересованных" анализаторов. На рис. 50 представлено действие дифференцировочного раздражения (звуковых щелчков, следующих в ритме 20 в 1 сек., их интенсивность 63 дб), которое вызывает отчетливо выраженное нарастание амплитуд колебаний и следование ритму раздражения в слуховой зоне коры и не вызывает отчетливого изменения электрической активности в остальных структурах ствола и коры мозга собаки № 3. Лишь в первую секунду действия звука имеет место непродолжительный (1,2 сек) эффект десинхронизации, совпадающий с высокоамплитудным отклонением в зрительной зоне коры. Весь же период действия звукового сигнала высокоамплитудные колебания в коре латеральной извилины отсутствуют, но они появляются в четко выраженной форме сразу же после окончания действия звукового сигнала. На рис. 26 были приведены ЭЭГ той же собаки, записанные в момент применения звукового раздражения более слабой интенсивности (тон 500 гц 49 дб), но являющегося сигналом условной реакции. Отчетливо видно множество волн типа вызванных потенциалов в зрительной зоне коры головного мозга.
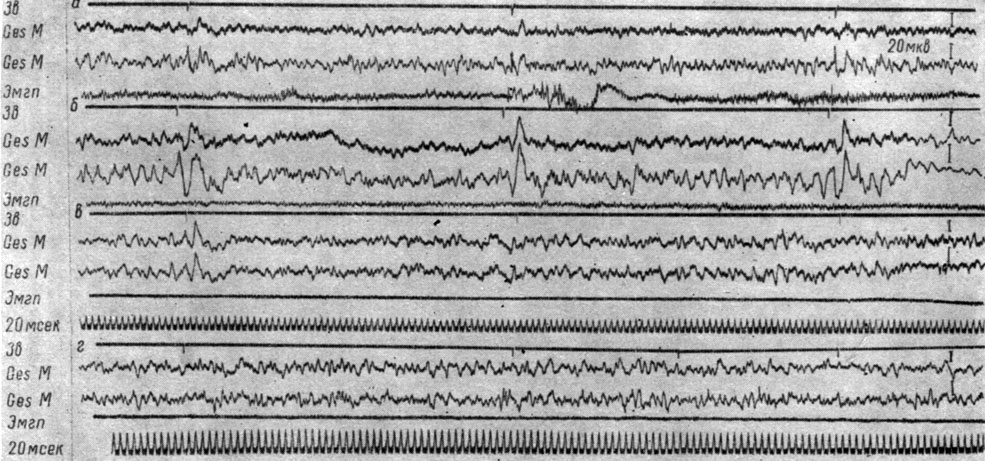
Рис. 50. Изменения ЭГ коры большого мозга и структур продолговатого мозга собаки № 3, наступающие при осуществлении дифференцировочного торможения на 18-е применение щелчков (20 в 1 сек, 63 дб, опыт № 73 от 20/V 1959 г.)
Детальный анализ параметров первичных вызванных ответов, появляющихся в корковом конце сигнального анализатора при действии дифференцировочного раздражения (звуковых щелчков в ритме 1 в 1 сек), показал, что характер их изменений зависит от этапа формирования дифференцировочного торможения, а также от порядкового номера щелчка в серии, служащей сигналом тормозной реакции.
Изучение параметров вызванных ответов на включение дифференцировочного стимула, т. е. на первый щелчок, показало, что они претерпевают в процессе образования и упрочения отрицательной реакции те же изменения, что и при формировании условной реакции, а именно, по мере выработки дифференцировки происходит сначала возрастание, а затем постепенное снижение амплитуды первой позитивной, негативной и второй позитивной компоненты ответа.
Различия между положительными и отрицательными сигналами выражаются лишь в динамике ответов на последующие щелчки. Сказанное иллюстрируют рис. 51 и 52. На рис. 51 представлены ЭЭГ эктосильвиевой извилины у собаки № 19 в разные периоды формирования дифференцировочного торможения. Видно, что величина вызванного ответа на первый щелчок после 10 применений тормозного сигнала (см. рис. 51, б) значительно превышает величину аналогичного ответа при первых применениях этого раздражителя (см. рис. 51, а). В ответ на включение сигнала после 70 его применений регистрируются ответы значительно меньшей амплитуды (см. рис. 51, в). В дальнейшем, когда число применений дифференцировочного стимула достигает 190-250 и больше, вызванные ответы перестают регистрироваться (см. рис. 51, г). Исчезновение вызванных потенциалов на действие первого щелчка в серии, служащей сигналом дифференцировочного торможения, происходит в тот же период, когда перестают регистрироваться вызванные потенциалы и на действие положительных сигналов оборонительной реакции.
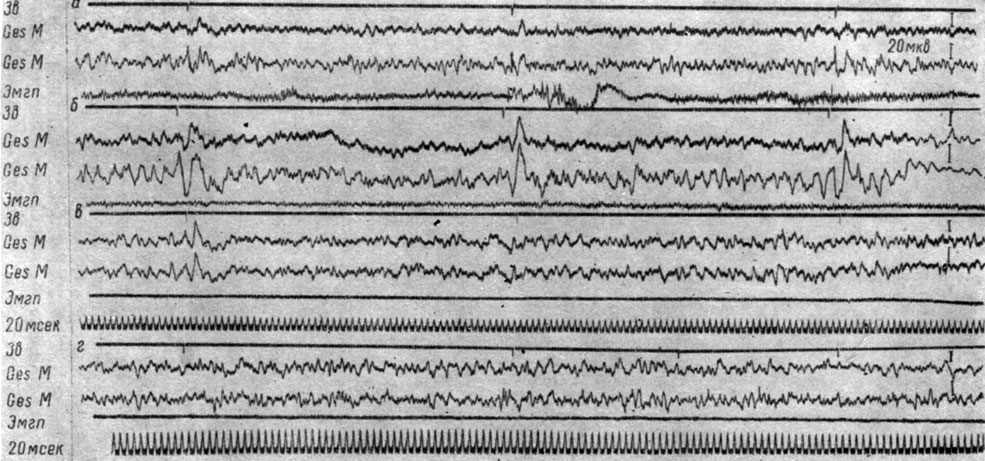
Рис. 51. Изменения вызванных потенциалов в разных участках первичной проекционной корковой слуховой зоны собаки № 19 при формировании дифференцировочного торможения на ритмические (1 в 1 сек) звуковые раздражения (щелчки 56 дб). а - 7-е применение (опыт 18 от 11 /VI 1964 г.); б - 51-е применение (опыт № 29 от 26/VI 1964 г.); в - 233-е применение (опыт № 95 от 8/XII 1964 г.); г - 251-е применение (опыт № 100 от 16/ХII 1964 г. - по T. С. Наумовой и Н. С. Поповой, 1967а)
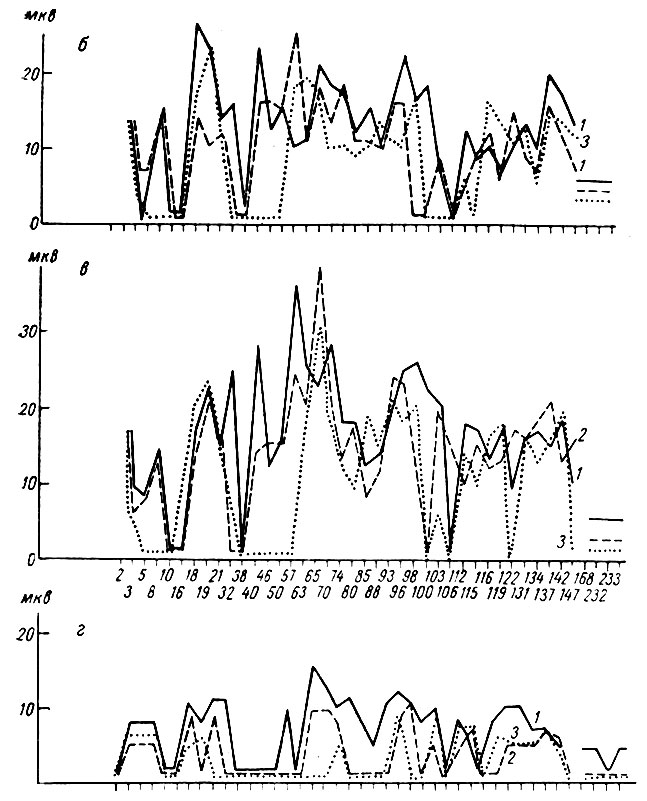
Рис. 52. Динамика амплитуды разных фаз первичных вызванных потенциалов при выработке дифференцировки на щелчки 1 в сек, 56 дб (собака № 19). Обозначается б, в, г, те же, что на рис. 22. 1 - амплитуда ответа на 1-й щелчок; 2 - на 2-й щелчок и 3 - на 3-й щелчок (по Т. С. Наумовой и Н. С. Поповой, 1967в)
На рис. 52 представлены результаты вычисления величины I позитивной, негативной и II позитивной компенент вызванных ответов центральной территории коркового конца слухового анализатора на первый (непрерывная линия), второй (пунктирная линия) и третий (линия точек) щелчки в серии, служащей сигналом дифференцировки. Видно, что у этой собаки возрастание величины первых трех компонент вызванного ответа на первый щелчок наступало после 10 применений дифференцировочного стимула и длилось до 70 проб, после чего амплитудная компонента вызванного ответа волнообразно начала уменьшаться, вплоть до полного исчезновения вызванных потенциалов, которое наступило после 250 применений тормозного раздражителя.
Сопоставление величины вызванных ответов на первый и последующие два щелчка показало, что в период увеличения этих ответов на первый щелчок происходит увеличение их (но менее выраженное) и на последующие щелчки (см. рис. 51, б и 52) подобно тому, что имело место при применении условных положительных сигналов. В период снижения амплитуды ответов на первый щелчок уменьшается величина вызванных потенциалов на второй и третий щелчки. К концу последнего периода, незадолго до исчезновения вызванных потенциалов, наступает момент, когда вызванные ответы появляются только на первый щелчок, на следующие его действия они отсутствуют (см. рис. 51, б). Иногда удается заметить, что сначала исчезают ответы на третий щелчок, а затем на второй и, наконец, первый. Так, например, у той же собаки № 19 реакция на третий щелчок перестала регистрироваться после 178-го применения тормозного сигнала, на второй и третий - после 221-й его пробы.
Все сказанное в отношении динамики амплитуды ответов на первый и последующие щелчки касается всех трех компонент вызванного потенциала (см. рис. 52).
В соответствии с двумя фазами изменений амплитудной характеристики вызванных ответов менялись их латентный период и длительность. Латентный период начала вызванного ответа, время до максимума позитивной и время до максимума негативной фаз ответа в первый период укорачивается, а затем постепенно удлиняется. Сказанное иллюстрирует рис. 53, построенный на основании измерений латентного периода компонент вызванного ответа на три щелчка, служащих дифференцировочным сигналом, у собаки № 19. Ответы на второй и третий щелчки (пунктирная линия и линия точек) наступают при выработке дифференцировочного торможения с тем же, большим или меньшим латентным периодом по сравнению с ответом на первый щелчок.
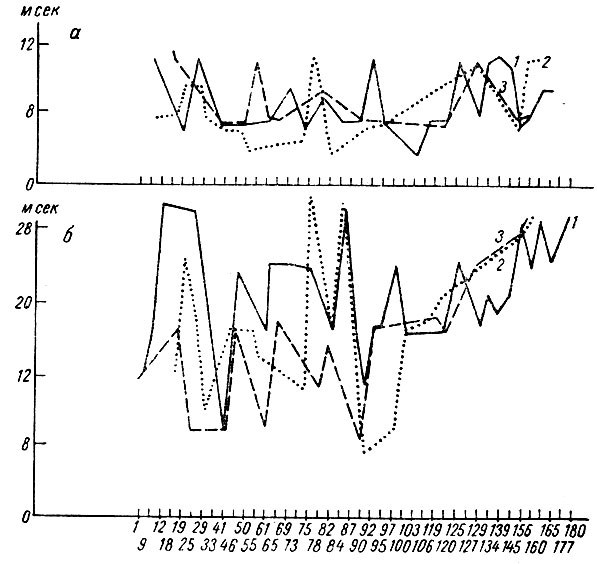
Рис. 53. Динамика латентного периода первичных вызванных потенциалов при выработке дифференцировки на щелчки 1 в сек, 56 дб (собака № 19). Обозначения а, б, в те же, что на рис. 23. Обозначения 1, 2, 3 те же, что на рис. 52 (по Т. С. Наумовой и Н. С. Поповой, 1967в)
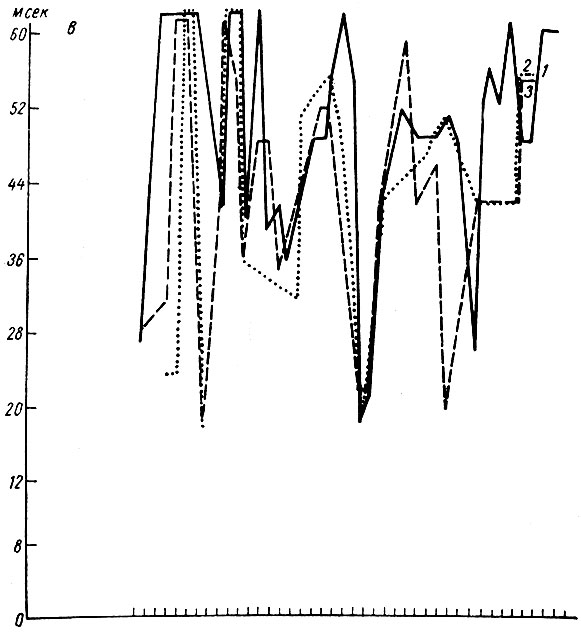
Рис. 53. Динамика латентного периода первичных вызванных потенциалов при выработке дифференцировки на щелчки 1 в сек, 56 дб (собака № 19). Обозначения а, б, в те же, что на рис. 23. Обозначения 1, 2, 3 те же, что на рис. 52 (по Т. С. Наумовой и Н. С. Поповой, 1967в)
Что касается длительности трех компонент вызванных потенциалов, то она сначала нарастает (вместе с увеличением амплитуды ответа) и снижается на фоне снижения их величины (рис. 54). Как видно на графике, снижение величины вызванных потенциалов к концу выработки рефлекса сопровождается вновь увеличением длительности ответов, которые после этого исчезают.
Рис. 54. Динамика длительности разных фаз первичных вызванных потенциалов при выработке дифференцировки на щелчки 1 в сек, 56 дб (собака № 19). Обозначения а, б, в те же, что на рис. 24. Обозначения 1, 2, 3 те же, что на рис. 52. (по Т. С. Наумовой и Н. С. Поповой, 1967в)
Итак, при выработке дифференцировочного торможения наступают сдвиги ЭГ, последовательно повторяющие все этапы формирования условных рефлексов, а затем применения дифференцировочного стимула не сопровождаются видимыми изменениями электрической активности. Отсутствие заметных изменений ЭГ касается всех изученных образований мозга: кохлеарных ядер, ядер нежного и клиновидного канатиков, внутреннего коленчатого тела, задних бугорков четверохолмия, ядер ретикулярной формации ствола мозга, территорий коры и т. д., т. е. как системы взаимодействующих анализаторов, так и других, условно говоря, межанализаторных зон.
Если выработка дифференцировочного торможения происходила на непрерывные звуковые сигналы, которые до выработки рефлексов вызывали, несмотря на многократные применения, выраженные сдвиги электрической активности (депрессию электрических процессов в стволовых отделах мозга, а также эффект десинхронизации электрических колебаний при отведении ЭЭГ от территорий коры накладными электродами), на всех этапах формирования отрицательной реакции на такие звуковые стимулы, по интенсивности значительно превышающие пороговые значения, превалировали процессы депрессии электрической активности в стволе и десинхронизация ее в коре головного мозга. Однако и в случае действия надпороговых тормозных раздражений последние переставали, в конце концов, сопровождаться видимыми сдвигами ЭЭГ.
Сопоставление электрографических картин активности, сопровождающих тормозные реакции на звуковые сигналы разной интенсивности, показывает, что эффект депрессии электрических потенциалов в ответ на действие достаточно интенсивных дифференцировочных стимулов в период еще недостаточно упроченного отрицательного рефлекса определяется физиологическими свойствами сильного звукового раздражения.
Если формирование дифференцировочного торможения происходило на интенсивные ритмические (1 в 1 сек) раздражения (щелчки), описанным ЭГ сопутствовали изменения параметров вызванных ответов в сигнальном анализаторе, которые зависели от этапа формирования дифференцировочного торможения, а также от порядкового номера щелчка в серии, служащей сигналом тормозной реакции. Изучение динамики амплитудной характеристики, латентного периода и длительности первых трех компонент вызванного ответа на включение дифференцировочного стимула, т. е. на первый щелчок, показало, что они претерпевают в процессе образования и упрочения отрицательной реакции те же изменения, что и при формировании условной реакции, а именно, по мере выработки дифференцировки происходит сначала возрастание, а затем постепенное снижение амплитуды первой позитивной, негативной и второй позитивной компоненты ответа. После этого ответы исчезают. В соответствии с двумя фазами изменений амплитудной характеристики вызванных ответов меняется их латентный период и длительность. Латентный период начала вызванного ответа, время до максимума позитивной и время до максимума негативной фазы ответа на все три щелчка в первый период укорачиваются, а затем постепенно удлиняются, возвращаясь к исходным величинам.
Что касается длительности компонент ответов на щелчки, то они увеличиваются с увеличением амплитуды тех же компонент ответов, а затем снижаются вместе с уменьшением последних. Перед исчезновением вызванных потенциалов происходит уменьшение их величины и одновременное возрастание их длительности.
Таким образом, включение дифференцировочного сигнала на всех этапах формирования этого вида торможения сопровождается теми же изменениями параметров вызванных ответов, что и включение сигнала оборонительной реакции.
В период формирования тормозной реакции амплитудная характеристика, латентный период и длительность вызванных потенциалов на следующие щелчки в серии, служащей дифференцировочным сигналом, претерпевает те же изменения, что и ответ на первый щелчок.
Вызванные потенциалы на второй и третий щелчки исчезают ранее ответа на первый щелчок. В конечном итоге, перестают регистрироваться и вызванные ответы на первый щелчок. Последнее происходит тогда, когда исчезают вызванные ответы из системы условнорефлекторного анализатора и при действии сигналов оборонительной реакции.
Как было показано, электрографическое выражение дифференцировочного торможения зависит от стадии его выработки, а также от силовых свойств стимула, на который формируется тормозная реакция. Выделено два периода образования дифференцировки: в первом периоде дифференцировочный звуковой раздражитель вызывает сдвиги ЭЭГ, последовательно повторяющие все этапы формирования условных рефлексов, вплоть до того, когда была введена дифференцировка. Если дифференцировочный стимул имеет ритмический характер, его введение приводит к увеличению амплитуды первичных ответов как на серию щелчков, служащих сигналом оборонительной реакции, так и на тормозные ритмические звуковые стимулы. Возрастание амплитуды вызванных потенциалов происходит одновременно с укорочением латентного периода ответов, что свидетельствует о повышении возбудимости сигнального анализатора к восприятию положительного и отрицательного условного раздражения в период формирования дифференцировки, т. е. в период активного анализа и дифференцирования раздражителей.
Эти наблюдения согласуются с данными авторов, утверждающих, что при выработке дифференцировки, так же как при формировании положительной реакции, имеет место возбуждение (К. С. Абуладзе, 1963; Р. С. Мнухина, 1963, и др.), сопровождающееся снижением порога возбудимости сигнального анализатора (У. Г. Гасанов, 1955).
Во втором периоде - периоде упрочения реакции - дифференцировочный сигнал начинает сопровождаться теми изменениями ЭЭГ, которые наблюдались при действии стимула до выработки рефлекса, а именно, выработанные тормозные реакции на звуковые раздражения подпороговой и пороговой интенсивности (в смысле появления электрографической реакции) не вызывают видимых изменений электрических потенциалов коры и подкорково-стволовых структур мозга, в том числе тех, которые входят в систему взаимодействующих анализаторов. Те же реакции на звуковые раздражения, значительно превышающие пороговые значения, длительный период времени выражаются в генерализованной депрессии электрических потенциалов всех исследованных систем мозга. Только после этого наступит период, в который и действие отрицательного сигнала надпороговой интенсивности перестает вызывать видимые изменения ЭЭГ. Таким образом, для более интенсивных стимулов исчезновение изменений электрической активности происходит значительно позднее, чем для подпороговых и пороговых раздражений. Если дифференцировочный сигнал носит ритмический характер, то его действие в период сформировавшейся дифференцировки сопровождается постепенно уменьшающимися вызванными потенциалами так, что, в конечном итоге, при действии дифференцировочного ритмического стимула наступает ответ на его включение, т. е. первый щелчок, а затем и эта реакция исчезает. Отмеченные изменения вызванных ответов касаются всех его компонент. В отличие от данных, полученных Р. С. Мнухиной (1963) на кроликах, в наших опытах не удалось обнаружить периода выработки дифференцировки, в которой регистрировалась бы лишь негативная компонента вызванного ответа.
Полученные нами факты позволяют заключить, что динамика изменений основных нервных процессов, развивающихся при применении дифференцировочных стимулов, не может рассматриваться вне связи с силовыми свойствами действующего раздражения. Как уже говорилось, всякое условное раздражение несет в себе, с одной стороны, свойства раздражителя как такового, с другой же стороны, свойства раздражителя как сигнала какой-то реакции. Если торможение формируется на физиологически более слабые раздражения, то реакция на действие сигнала как такового обычно угасает еще до выработки тормозного рефлекса, и его действие заключает в себе свойства преимущественно сигнального значения раздражителя.
При выработке тормозных рефлексов на физиологически более сильные (надпороговые) звуковые раздражения электрографическую реакцию на них не удается угасить даже при многократных повторениях, раздражителя, достаточных для угашения реакций на более слабые звуки. Таким образом, выработка рефлекса происходит на звуковые стимулы, которые оказывают влияние и как таковые, и как сигнальные, поэтому обычно условнорефлекторные изменения электрической активности мозга в ответ на воздействие физиологически сильных сигналов, в первую очередь, выражаются в реакции на действие самого раздражителя, а именно, депрессии колебаний электрических потенциалов, наблюдавшейся и до выработки рефлекса. Этим объясняется и значительное сходство изменений, сопровождающих положительные и тормозные реакции на сигналы значительной интенсивности.
В случае действия ритмического раздражения это выражается в значительном сходстве вызванных первичных ответов, сопровождающих начальные периоды действия положительного и тормозного сигналов.
Отсутствие существенных изменений электрической активности при действии тормозных сигналов в период упроченной дифференцировки, а также появление эффекта депрессии электрических потенциалов в период ее формирования на стимулы, по интенсивности значительно превышающие пороговые значения, согласуются с данными других авторов (Г. Т. Сахиулина, 1957; О. В. Верзилова, 1958; Gluck, Rowland, 1959; Rowland, 1960; Ю. Г. Кратин, 1960, 1966; Р. С. Мнухина, 1963; Ю. Г. Кратин, В. Н. Андреева, 1966), обнаруживших эффект десинхронизации или отсутствия каких-либо сдвигов ЭЭГ при развитии дифференцировочного торможения у животных и человека. Отмечена зависимость электрографического выражения тормозной реакции и от сложности дифференцировки. Так, Ю. Г. Кратиным (1960) показано, что отчетливый эффект десинхронизации в коре мозга человека можно было видеть при развитии дифференцировочного торможения в том случае, когда различие между положительным и дифференцируемым сигналом было очень малым. Если дифференцировке подвергались звуковые сигналы, значительно отличающиеся от положительных, дифференцируемый звук не сопровождался никакими видимыми изменениями ЭЭГ. Очевидно, при формировании отрицательной реакции на сигналы, по интенсивности значительно превышающие пороговые значения, а также в условиях трудной дифференцировки происходит усиление ориентировочного рефлекса, который сопровождается повышением тонуса структур мозга и депрессией его электрической активности (Е. Н. Соколов, 1958, 1959, 1960).
При развитии дифференцировочного торможения нам не удалось наблюдать медленных изменений электрической активности, которые имели место в ряде исследований ЭЭГ собак и кошек, осуществляемых на базе оборонительной (М. Я. Рабинович, 1958, Roy и сотр., 1961, и др.) и пищевой (Н. Н. Любимов, 1960) методик.
При упроченной дифференцировке на звуковые сигналы нами не наблюдалось четких изменений электрической активности структур слухового анализатора и в том числе первичных переключательных реле (кохлеарных ядер). Можно думать, что уже на этом уровне восприятие сигнала тормозной реакции происходит по-иному, чем положительного сигнала. Поэтому нельзя не признать справедливой точку зрения И. П. Павлова и тех его последователей, которые говорят о связи торможения с процессами, происходящими в первую очередь в анализаторе условного сигнала (Л. Г. Воронин, 1952; А. А. Травина, 1954; П. С. Купалов, 1960; А. И. Ройтбак, 1960; У. Г. Гасанов, 1962, и мн. др.). Речь должна идти о сдвигах функционального состояния в компонентах всего анализатора, воспринимающего условный сигнал, начиная с первичных переключательных его реле.
Процесс, несомненно, начинает формироваться в анализаторе, воспринимающем отрицательный сигнал, но этим дело не ограничивается, потому что исчезают и те процессы активации сенсомоторной зоны коры, которые обычно сопутствуют условному оборонительному рефлексу. Это явление свидетельствует о том, что формирование дифференцировки связано и с процессами в эффекторных корковых компонентах реакции, с деятельностью которой связана реализация условного рефлекса. В этом отношении нужно согласиться и с И. Я. Перельцвейгом (1907), Н. А. Кашерининовой (1908), П. К. Анохиным (1932, 1958а), Н. А. Рокотовой (1953), И. И. Короткиным и М. М. Сусловой (1955), Б. И. Ходоровым (1955), Г. В. Скипиным (1956), Р. Г. Зевальдом (1958), М. А. Панкратовым (1960) и др., утверждающими, что развитие внутреннего торможения связано с процессами в эфферентной части дуги условного рефлекса.
Представленный материал позволяет утверждать, что как при формировании условного рефлекса, так и при образовании дифференцировки участвует констелляция структур мозга, поэтому трудно думать, что процесс обратного развития временной связи идет только в одной какой-то (афферентной или эфферентной) системе. Он охватывает как системы взаимодействующих анализаторов (И. П. Павлов; П. С. Купалов, А. М. Ушакова, 1931; А. А. Травина, 1954; П. С. Купалов, 1960) и те пути, которые их соединяют (Э. А. Асратян, 1941, 1955, 1959), так и структуры, так или иначе причастные к этой реакции.
Начальный момент действия отрицательного стимула на всех этапах формирования дифференцировки сопровождается мгновенным усилением активности, которое регистрируется в системе структур, образующих на каждом данном этапе образования условного рефлекса доминантную констелляцию. Так как и включение ритмического дифференцировочного раздражения (первого щелчка) вызывает те же изменения, что и действие первого щелчка в серии их, служащей сигналом оборонительной реакции, то и в этом смысле начальные эффекты положительного и тормозного стимулов ничем по существу не отличаются друг от друга. Эти наблюдения говорят о том, что сигнал дифференцировки всегда проходит анализаторную обработку, прежде чем вызвать характерную для него реакцию, т. е. также связан с распространением опережающего возбуждения (П. К. Анохин, 1962).
Лишь в период, следующий после включения раздражителя в стадии упроченной дифференцировки не происходит никаких видимых изменений ЭЭГ: ни суммарных сдвигов активности, ни вызванных потенциалов.
Отсутствие изменений электрической активности в разных звеньях слухового анализатора при дальнейшем действии дифференцировочного сигнала как подпороговой и пороговой, так и надпороговой интенсивности свидетельствует о том, что состояние возбудимости слухового анализатора на протяжении действия дифференцировочного сигнала снижается по сравнению с состоянием в начальный момент действия стимула. Об этом же говорят и исследования параметров вызванных потенциалов. Перед исчезновением ответов на 3, 2, а затем и 1 щелчки происходит падение их величины и увеличение длительности. Описываемые сдвиги касаются всех компонент ответа, т. е. позитивного, негативного и второго позитивного колебания. Снижение возбудимости начинается с конца раздражения, сначала на 3-й щелчок, а затем уже на 2-й и, наконец, на 1-й.
Иными словами, по мере развития дифференцировочного торможения происходит обратное развитие тех процессов, которые формируются при образовании условного рефлекса, т. е. снижение уровня возбудимости в первую очередь сигнального анализатора, достигнутого в процессе формирования условной реакции (А. А. Князева, И. Э. Барбель, 1952; А. А. Гюрджан, 1953; А. М. Марусева, Л. А. Чистович, 1954; Е. Н. Соколов, 1959, и др.).
Снижение возбудимости сигнального анализатора в процессе действия дифференцировочного раздражителя в период упроченной тормозной реакции может быть констатировано лишь по отношению к тому уровню, который был достигнут в процессе переноса на дифференцировочный раздражитель свойств условного сигнала (в начале выработки дифференцировки). Но понижается ли возбудимость сигнального анализатора в процессе применения дифференцировочного раздражителя ниже исходной величины, т. е. появляются ли в нем сдвиги функционального состояния, которые могли бы быть расценены как признаки торможения? Приведенные выше факты дают право думать, что состояние условнорефлекторного анализатора в период осуществления тормозной реакции в стадии сформировавшейся дифференцировки не снижается ниже исходного уровня, так как латентный период и длительность ответов обычно не превышают те же показатели вызванных потенциалов до выработки на них тормозной реакции. Изменение этих параметров ответов наступает лишь в тот период, когда и на действие условных сигналов оборонительной реакции происходит постепенное уменьшение величины, увеличение латентного периода и длительности всех компонент первичного ответа, наступающее перед самым исчезновением вызванных потенциалов. Не выявляется электрографических признаков торможения и по показателям суммарной электрической активности.
Таким образом, при действии дифференцировочных раздражений в сигнальном анализаторе не выявляется никаких признаков торможения. В этом смысле наши данные согласуются с наблюдениями А. А. Гюрджан (1953), который показал, что при действии дифференцировочного светового раздражения (так же как при действии индифферентных раздражений) граница поля зрения остается неизменной, в то время как при применении условного сигнала она резко возрастает.
Не обнаружено было нами признаков торможения и в других системах мозга и, в частности, стволовых и кортикальных компонентах кожно-двигательного анализатора. Как в этой территории, так и в других зонах мозга (стволово-подкорковых и кортикальных) при развитии дифференцировочного торможения обнаруживались те изменения ЭЭГ, которые сопровождали действие того же раздражителя до выработки на него отрицательной реакции и которые зависели от силовых свойств стимула.
Не найдено было признаков торможения и в ретикулярной формации, которой (Gastaut, 1955; А. Гасто и А. Роже, 1958) приписывают роль главного участника дифференцировочного торможения. Дифференцировочное торможение, по мнению этих авторов, связано с торможением таламических неспецифических систем, так как при прочно выработанных дифференцировках, по данным этих исследователей, наблюдается локальная синхронизация в безусловнорефлекторной корковой зоне.
Несомненно, ретикулярные структуры являются участниками тормозных процессов в той же мере, в какой они участвуют в формировании условных рефлексов, но приписывать им роль инициаторов торможения нет никаких оснований.
Все это свидетельствует, по-видимому, о том, что дифференцировочное торможение оборонительных рефлексов не сопровождается процессами торможения в собственном смысле слова.
Нам представляется справедливой точка зрения тех авторов, которые считают, что механизмы, лежащие в основе дифференцирования, формируемого на базе оборонительных условных рефлексов, будут иными, чем пищевых (П. К. Анохин, 1958а; А. И. Ройтбак, 1959). В основе дифференцировки оборонительных рефлексов, по-видимому, не лежит процесс торможения в истинном смысле этого слова или развитие трудного состояния (доминанты другого рода) (П. К. Анохин, 1958а), как в случае торможения пищевых рефлексов. В случае оборонительных рефлексов речь идет скорее о своеобразном угашении условной реакции, когда отрицательный раздражитель превращается в индифферентный стимул. По-видимому, при формировании дифференцировки происходит снижение реактивности сигнального анализатора, которое наступает вследствие отсутствия притока неспецифической для него болевой афферентации, формировавшей избирательную его восприимчивость к условным раздражителям. Снижение уровня возбуждения условного анализатора сопровождается уменьшением потока слуховой информации в направлении эффекторных компонент оборонительного условного рефлекса, что, в свою очередь, ведет к распаду всей системы временной связи.
Постепенное обратное развитие (И. С. Бериташвили, 1960) условного рефлекса происходит, таким образом, под контролем обратных связей (от двигательного анализатора к слуховому), которые в свою очередь находятся под влиянием поступательных связей (от слухового анализатора к двигательному). Отсутствие подкрепления ведет к обратному развитию как поступательных, так и обратных путей рефлекса, хотя возможно, что развитие последних и превалирует (Н. С. Бериташвили, 1960).
Наши данные свидетельствуют о том, что обратному развитию подвергается вся констелляция нервных центров, участвующая в формировании условной реакции. Очевидно, это касается как системы взаимодействующих анализаторов, так и структур, участвующих в целостной реакции организма. Весь комплекс целостной реакции организма со всеми его вегетативными компонентами на более сильные раздражения бывает иным, чем на раздражения меньшей интенсивности, поэтому изменения функционального состояния будут наступать в одних случаях в меньшем, а в других случаях в большем количестве образований мозга, но всегда речь будет идти о дезинтеграции процессов во всех нервных центрах, ранее участвовавших в формировании условного оборонительного рефлекса.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'