Особенности взаимодействия специфической и неспецифической афферентации, зависящей от силовых свойств сочетаемых раздражений
Исследование закономерностей распространения возбуждения в центральной нервной системе началось еще в прошлом веке. Так, Pfluger (1853) показал, что при усилении болевых раздражений на смену реакциям эффектора, соответствующего данной рефлексогенной зоне, приходят реакции, определяющиеся процессами иррадиации возбуждения сначала в одноименной, а затем и в противоположной половине спинного мозга. В дальнейшем многочисленные эксперименты И. М. Сеченова (1889-1890), Sherrington (1906) и других показали значение силы внешних раздражений для объема иррадиации возбуждений.
Зависимость эффекта от силовых характеристик раздражителей была обнаружена и в исследованиях закономерностей высшей нервной деятельности.
В работах школы И. П. Павлова было показано, что характер иррадиации и концентрации процессов находится в зависимости (при прочих равных условиях) от интенсивности действующих сигналов. На основании опытов В. В. Рикман (1928) с растормаживанием тормозной фазы и торможением возбудительной фазы запаздывающего рефлекса при помощи раздражителей различной силы И. П. Павлов в 1932 г. сформулировал закон иррадиации и концентрации возбуждения и торможения в зависимости от силы раздражителя; "при слабом напряжении этих процессов они с первого момента и с места их возникновения иррадиируют, при достаточно сильном - концентрируются и при чрезвычайно сильном - опять иррадиируют" (И. П. Павлов, 1949, т. III, стр. 370).
Первоначально считалось, что процессы иррадиации и концентрации, сопровождающие высшую нервную деятельность, складываются в коре головного мозга, однако данные И. Я. Перельцвейга (1907), П. К. Анохина (1932), Э. А. Асратяна (1934), А. М. Павловой (1940) дали возможность считать, что эти процессы захватывают и безусловные центры.
В настоящее время исследования закономерностей движения и взаимодействия нервных процессов электрофизиологическим способом проводятся без достаточного учета фактора интенсивности действующих раздражений, хотя отдельные работы в этой области и имеются (Motokawa, 1954; Г. В. Гершуни, 1955; Petsch, Stumf, 1960, и др.).
Как отмечалось в главе II, внешние стимулы, действующие на собак, находящихся в экспериментальной комнате в состоянии бодрствования, вызывают изменения ЭЭГ коры и структур ствола мозга, которые зависят не только от функционального состояния животного и индивидуальных особенностей его нервной системы, модальности, но также от интенсивности действующего раздражения (см. рис. 7).
Как указывалось, при формировании условной связи выступают закономерные изменения электрических процессов, которые являются характерными для каждого этапа выработки оборонительного рефлекса. Вместе с тем, удалось обнаружить, что хотя эта зависимость и имеет место, тем не менее электрографические картины активности, сопровождающие осуществление условных оборонительных рефлексов, зависят не только от индивидуальных особенностей нервной системы подопытных животных и функционального состояния их нервной системы, но также от силовых свойств сочетаемых раздражителей.
Как известно, изменения ЭГ, регистрируемые на субкортикальном и кортикальном уровнях при биполярном способе их отведения погружными электродами, которые наблюдаются на первых этапах формирования оборонительного условного рефлекса, имеют генерализованный характер, по мере же сочетаний они становятся более локальными. Локальные процессы бывают отчетливо выражены при применении звуковых сигналов подпороговой и пороговой интенсивности (тон 500 гц 30-50 дб), которые до выработки рефлекса не вызывают существенных изменений ЭЭГ.
Следует указать, во-первых, на особенности электрографических реакций, которые сопровождают формирование условных оборонительных рефлексов на подпороговые звуковые раздражения. У тех животных, у которых образование рефлекса на звуки пороговой интенсивности в I стадии сопровождалось генерализованной депрессией колебаний электрических потенциалов, выработке рефлекса на сигналы подпороговой интенсивности с самого начала сопутствовал эффект диффузной экзальтации, характерный для второго периода генерализации. На рис. 44, а представлены изменения ЭГ собаки № 4, сопровождающие выработку условных рефлексов на относительно слабые тоновые раздражения (500 гц 40 дб). Формирование рефлекса на эти раздражения происходило после образования рефлекса на более сильные (пороговые) сигналы (48 дб), фазы которого описаны были ранее (см. рис. 14, 25 и 31).
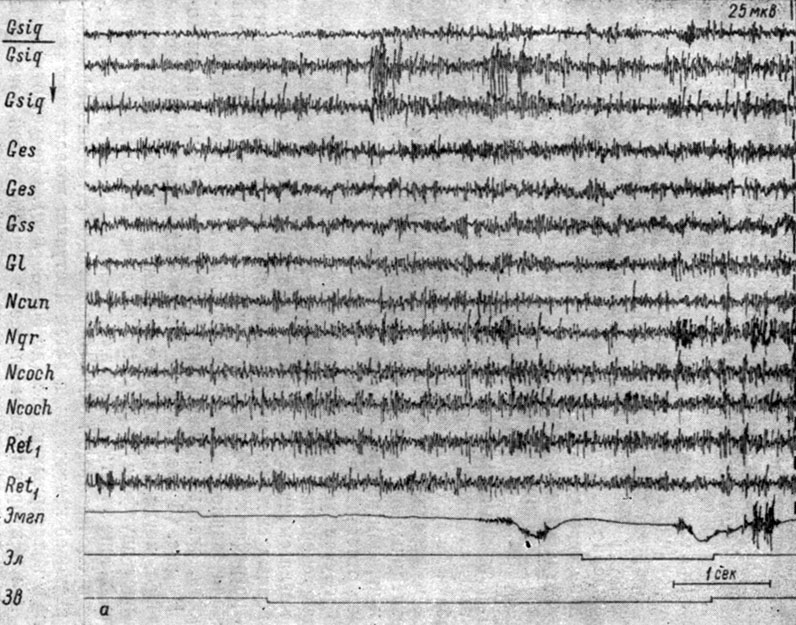
Рис. 44. Изменения ЭГ собаки № 4, наступающие во II (а) и III (б) стадиях формирования условного оборонительного рефлекса на подпороговые звуковые раздражения (тон 500 гц 40 дб). а - 2-е сочетание (опыт № 93 от 27/II 1959 г.); б - 9-е сочетание (опыт № 95 от 30/II 1959 г.- по Т. С. Наумовой, 1962а)
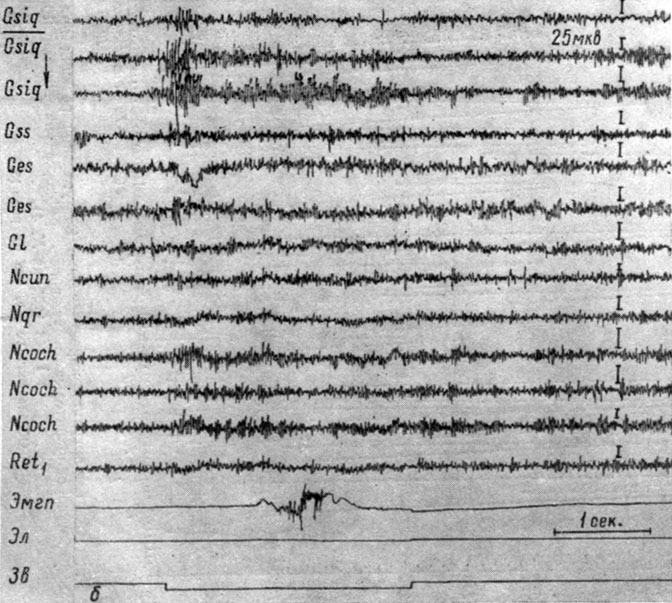
Рис. 44. Изменения ЭГ собаки № 4, наступающие во II (а) и III (б) стадиях формирования условного оборонительного рефлекса на подпороговые звуковые раздражения (тон 500 гц 40 дб). а - 2-е сочетание (опыт № 93 от 27/II 1959 г.); б - 9-е сочетание (опыт № 95 от 30/II 1959 г.- по Т. С. Наумовой, 1962а)
Перераспределение электрической активности в структурах ствола мозга в данном случае происходило значительно быстрее. Рефлекс обнаружился уже на 2-м сочетании (см. рис. 44, а) и выражался в генерализованном возрастании электрической активности структур продолговатого мозга и сенсомоторной зоны коры (фаза генерализованного снижения активности отсутствовала). На 9-м сочетании (см. рис. 44, б) наступила четко выраженная концентрация процессов в бульбарных звеньях слухового и кортикальных компонентах двигательного анализаторов, выражающаяся в более отчетливом возрастании электрической активности (почти втрое) в этих структурах. Вместе с тем, нельзя не отметить и некоторого роста активности в ретикулярных и других структурах ствола, зрительной и слуховой зонах коры.
При применении условных звуковых стимулов, по интенсивности значительно превышающих пороговые значения (тон 500 гц 60-100 дб; звуковые щелчки 56 дб; звонок и зуммер интенсивностью 700-100 дб), превалируют генерализованные сдвиги ЭЭГ, сходные с теми, которые вызывают эти раздражения до выработки рефлекса (депрессия активности), и локальные процессы выступают менее отчетливо. Как правило, формирование рефлекса на звуковые раздражения, превышающие по интенсивности пороговые значения, происходило после выработки условных рефлексов на звуковые сигналы подпороговой и пороговой интенсивности. При повторном образовании рефлекса, как известно, его появление происходит значительно быстрее. Прохождение электрографических стадий при повторном формировании временной связи также должно ускориться. Однако при становлении рефлекса на звуковые сигналы, по интенсивности значительно превышающие пороговые значения, прохождение I стадии генерализации (стадия генерализованной депрессии активности) значительно затягивается, в то время как другие стадии либо исчезают (II стадия генерализации - стадия генерализованной экзальтации активности), либо выступают неотчетливо (III стадия - первый период концентрации). Как видно на рис. 45, эффект генерализованной депрессии электрической активности на щелчки 10 в 1 сек (56 дб, у собаки № 4), охватывающий исследованные бульбарные структуры, в середине первой стадии становится менее глубоким в кохлеарных ядрах, в ядрах нежного и клиновидного канатиков и нижнем центральном ядре ретикулярной формации. В этот период отчетлива активация сенсомоторной зоны коры.
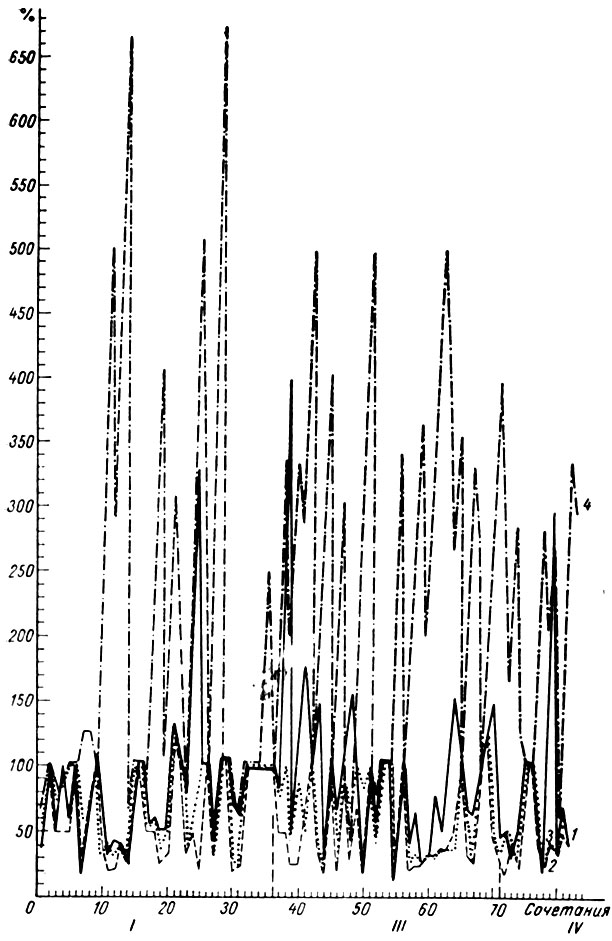
Рис. 45. Амплитудная характеристика ЭГ условной реакции при выработке оборонительного условного рефлекса на щелчки 10 в 1 сек (собака № 4). Обозначения те же, что на рис. 15
В случае условных сигналов надпороговой интенсивности стадия генерализованного снижения активности мозговых структур могла смениться стадией с более выраженными процессами в основных системах анализаторов лишь в форме двухфазного электрографического ответа, в котором I фаза представляла генерализованное снижение активности, а II - возрастание активности, охватывающее преимущественно системы анализаторов. На рис. 46 представлена третья стадия выработки того же рефлекса, когда надпороговое условное звуковое раздражение вызывает двухфазную реакцию: вначале наблюдается отчетливое снижение электрической активности (до 8-12 мкв) в структурах продолговатого мозга, совпадающее с условнорефлекторной реакцией конечности, а затем наступает возрастание активности в кохлеарных ядрах (до 30-35 мкв). Следует отметить, что, как и в предыдущем случае, в сенсомоторной зоне коры наступало возрастание активности (до 50 мкв), более выраженное при активации в кохлеарных ядрах. На рис. 45 видно, что эти изменения сопровождают все рефлексы вплоть до IV стадии. Отмеченные особенности наблюдаются как при исследовании бульбарных, так и других территорий мозга.
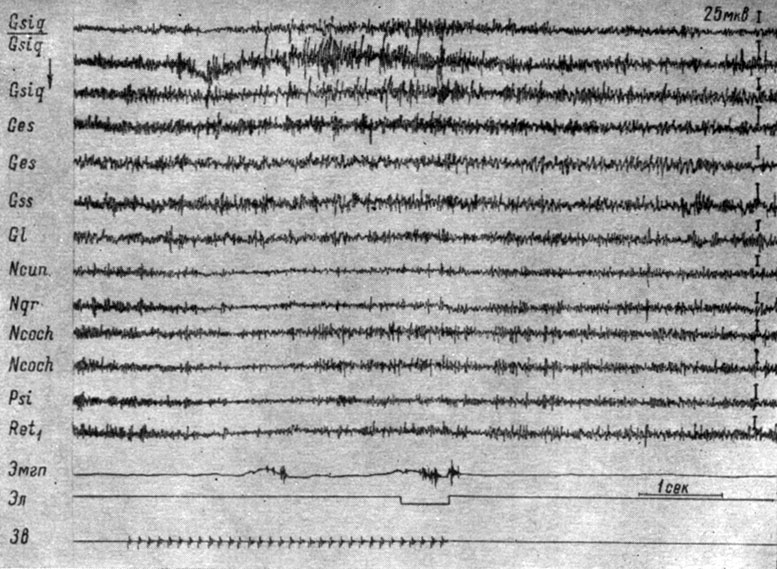
Рис. 46. Изменения ЭГ собаки № 4, наступающие в III стадии формирования условного оборонительного рефлекса на звуковое раздражение, значительно превышающее пороговые значения (щелчки 56 дб, 10 в 1 сек). 71-е сочетание (опыт № 83 от 29/V 1959 г. - по Т. С. Наумовой, 1962а)
Силовая характеристика звукового стимула сказывается и на величине вызванных потенциалов, которые находятся от нее в прямой зависимости. Факт этот общеизвестен.
Аналогичным образом различаются потенциалы по абсолютным значениям их амплитуды и при формировании временных связей на сигналы разной интенсивности. Вместе с тем, нам не удалось найти принципиальных различий в поведении вызванных ответов в первичной проекционной зоне слуха при выработке оборонительных условных рефлексов на звуковые сигналы пороговой и надпороговой интенсивности. Наблюдения Ван Тай-Ань и Н. И. Незлиной (1963) свидетельствуют о том, что динамика вызванных потенциалов и на сигналы подпороговой интенсивности существенно не отличается от динамики этих ответов на надпороговые стимулы. Однако тот факт, что изменения суммарной электрической активности при становлении рефлексов существенно зависят от силовых свойств условного сигнала, дает основание полагать, что и по динамике параметров вызванных ответов должны существовать различия в особенностях протекания каждой фазы выработки рефлекса на сигналы разной интенсивности. Разработка этого вопроса представляет большой интерес для выяснения особенностей иррадиации и концентрации основных нервных процессов в системе сигнального анализатора при формировании условных рефлексов.
Полученные нами данные позволяют заключить, что динамика изменений основных нервных процессов в формациях мозга при замыкании оборонительных условных рефлексов не может рассматриваться вне связи с интенсивностью действующего раздражения. Всякая условная стимуляция несет в себе, с одной стороны, свойства раздражителя как такового, с другой стороны, свойства раздражителя как сигнала какой-то реакции. Если происходит выработка рефлекса на звуки подпороговой или пороговой интенсивности, то до нее электроэнцефалографическая реакция обычно угашается. Вследствие этого угашается реакция на раздражение как таковое, и при дальнейшем приобретении этим раздражителем условнорефлекторного значения последнее заключает в себе свойства преимущественно его сигнального значения. Естественно, что при выработке рефлекса на раздражения подпороговой и пороговой интенсивности (в смысле вызова электрографической реакции) наблюдаются явления, в которых относительно слабо проявляются свойства самого раздражителя и выступает преимущественно условнорефлекторная окраска сигнала, поэтому действие подпороговых и пороговых стимулов вызывает изменения ЭЭГ, совершенно отличные от тех, которые наступают в ответ на эти же раздражения до выработки рефлекса.
Иное положение имеет место при выработке рефлекса на физиологически сильные звуковые раздражения, т. е. раздражения, значительно превышающие пороговые значения, которые оказывают влияние и как таковые, и как сигнальные. Вследствие этого условнорефлекторные изменения электрической активности мозга в ответ на воздействие сигналов, по интенсивности превышающих пороговые значения, в первую очередь выражаются в реакции на действие самого раздражителя, т. е. в появлении генерализованных изменений электрической активности, ее депрессии, которая маскирует собой локальные условнорефлекторные сдвиги ЭГ, появляющиеся в процессе формирования рефлекса. Отличие этой реакции от угнетения активности, наблюдавшегося в начальный период выработки рефлекса на звуковые раздражения пороговой интенсивности, состоит в том, что здесь эта реакция определяется физиологическим действием звука как такового, там же она происходила от сигнальных оборонительных свойств слабого звукового раздражения.
По мере упрочения реакции на интенсивные звуковые сигналы вслед за генерализованным снижением активности выступала известная концентрация процессов в слуховой системе и моторном представительстве реагирующей лапы. Тем не менее сама условнорефлекторная реакция осуществлялась на фоне генерализованной депрессии активности; иными словами, концентрация процессов во взаимодействующих анализаторах вуалировалась физиологическим действием звукового раздражения, которое в силу своих физических свойств оказывает более генерализованное влияние на центральную нервную систему, чем при действии раздражений подпороговой и пороговой силы.
Обращает на себя внимание тот факт, что как в случае действия подпороговых, пороговых, так и надпороговых раздражений изменения ЭЭГ в коре сенсомоторной зоны оказываются в значительной мере сходными, а именно в обоих случаях с помощью накладных электродов регистрируется возрастание частоты и амплитуд электрических колебаний или (у других собак) десинхронизация активности. Это свидетельствует о том, что процессы взаимодействия специфической и неспецифической афферентации в этой зоне мозга, определяющие замыкание, в обоих случаях сходны.
Таким образом, доминантная констелляция, формирующаяся на каждом этапе выработки условного оборонительного рефлекса, осуществляется при большем (при более сильных сигналах) или меньшем (при менее сильных сигналах) охвате структур мозга возбуждением.
Не менее важное значение для характера изменений электрической активности имеет и интенсивность болевого раздражения, используемого в качестве безусловного подкрепления. Как известно, сильное безусловное подкрепление резко увеличивает интенсивность слабого условного раздражителя, придавая ему особую значимость (П. С. Купалов, 1960; И. Б. Козловская, 1961). При использовании сильных (надпороговых) болевых подкреплений (электрических раздражений конечности) условнорефлекторные электрографические изменения обычно также приобретали генерализованный характер (генерализованное снижение электрической активности) независимо от интенсивности звука, на который вырабатывается условная реакция; поэтому в наших экспериментах в качестве подкрепления использовались пороговые электрокожные раздражения, которые вызывали легкий подъем правой передней лапы.
Таким образом, и силовая характеристика безусловнорефлекторного раздражения сказывается на характере условнорефлекторных изменений электрической активности мозговых структур с акцентом на диффузные процессы.
Исходя из сказанного, понятно, что те или иные особенности формирования электрических реакций можно уловить, лишь широко варьируя интенсивности сочетаемых раздражений.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'