Глава V. Некоторые особенности взаимодействия специфической и неспецифической афферентации в сигнальные и межсигнальные периоды
Индивидуальные особенности взаимодействия специфической и неспецифической афферентации в процессе осуществления безусловных и условных рефлексов
Характер сдвигов потенциалов головного мозга в большой мере зависит и от индивидуальных особенностей нервной системы подопытных животных. В школе И. П. Павлова типологические особенности высшей нервной деятельности связывают преимущественно со свойствами коры. Большинство электрофизиологических работ, проведенных в этом направлении, было посвящено сопоставлениям электрической активности коры и проявлений темперамента у человека (Gastaut и сотр., 1951, 1957; Mundy-Castle, 1957; Remond, Lesevre, 1957; Shipton, Walter, 1955; Bekkering и сотр., 1957; Дж. В. Н. де Ланге и сотр., 1958, и др.). Однако результаты этих сопоставлений представляются недостаточно убедительными из-за отсутствия строгих критериев характеристики темпераментов.
Сложность и разноречивость полученных данных (можно сослаться на выступление А. Гасто по докладу Дж. В. Н. де Ланге и сотр. на Московском коллоквиуме в 1958 г.), видимо, явились причиной значительного ослабления интереса к этой проблеме.
Сопоставлению электрических процессов коры у животных разного типа нервных систем посвящено довольно ограниченное количество исследований, в одних из которых сопоставлялись электрическая активность и темпераменты (Faure, Rougier, 1950; И. А. Пионтковский, А. М. Иваницкий, 1958), другие касались различий в электрических реакциях коры на безусловные и условные раздражения у собак разных типов высшей нервной деятельности (В. Д. Глезер и сотр., 1954).
Сложившееся в школе И. П. Павлова мнение, что типологические особенности высшей нервной деятельности, а следовательно, и индивидуальные свойства нервной системы определяются качествами клеточных элементов больших полушарий, исходит из положения о корковой локализации замыкания. Однако постоянно подчеркивавшаяся И. П. Павловым мысль о теснейшей взаимосвязи и взаимодействии коры и подкорки дает основание думать, что типологические и индивидуальные особенности определяются свойствами не только коры головного мозга, но нервной системы в целом. Эта мысль высказывалась в работах П. К. Анохина (1941), Б. В. Павлова и Н. А. Шустина (1948), Л. Г. Лейбсон и Т. Ф. Комаровой (1956).
В наших экспериментах обнаружено, что внешние раздражения, действующие на собак, находящихся в экспериментальной комнате в состоянии бодрствования, вызывают изменения ЭГ коры и структур ствола мозга, которые могут быть совершенно различными у разных животных, т. е. зависят от индивидуальных особенностей их нервной системы.
Сказанное касается реакций и на световые, и на звуковые, и на электрокожные раздражения, однако особенно четко индивидуальные различия проявляются в реакциях на болевые стимулы.
На рис. 43 представлены ЭГ двух зон коры и структур среднего мозга двух собак при действии порогового электрокожного раздражения на переднюю лапу. Отчетливо видно, что действие порогового электрокожного раздражения сопровождается у собаки № 9 (рис. 43, а) резким снижением (иногда до полного исчезновения) колебаний электрической активности стволовых структур и возрастанием ритма (с 40-45 до 50-55 в 1 сек) в записях ЭЭГ сенсомоторной и слуховой зон коры накладными электродами. Такие резкие колебания активности были наиболее демонстративными у 3 животных. У других собак (внешне характеризующихся спокойным, уравновешенным поведением, и таких в наших опытах было 6) колебания частотно-амплитудных показателей при тех же условиях были слабо выражены. На рис. 43, б видно, что у собаки № 21 в ответ на действие болевого стимула пороговой интенсивности происходит незначительное возрастание разности потенциалов как в структурах ствола, так и в сенсомоторной и слуховой корковых территориях. В двух демонстрировавшихся записях изменения ЭГ специфических и неспецифических структур ствола были сходными. Между этими крайними представителями имелись все переходные градации, и их мы наблюдали у 17 животных.
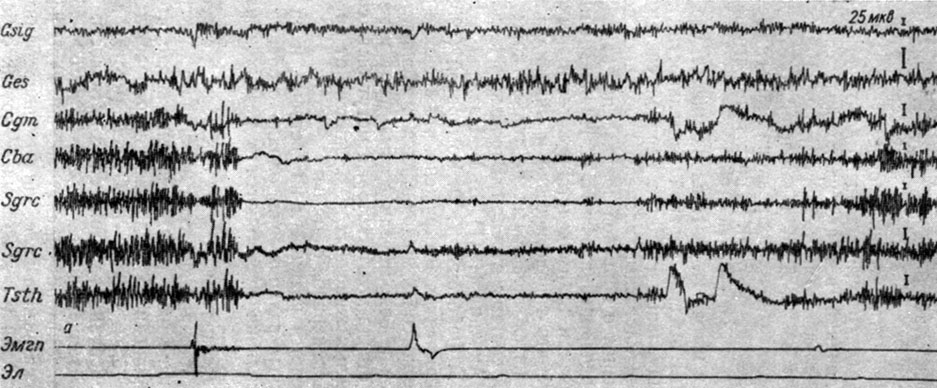
Рис. 43. Изменения ЭГ коры большого мозга и структур среднего мозга собаки № 9 (а) и собаки № 21 (б) при действии порогового электрического тока на правую переднюю лапу (а - опыт № 5 от 22/II 1958 г.; б - опыт № 12 от 12/IV 1958 г. - по Т. С. Наумовой, 1961)
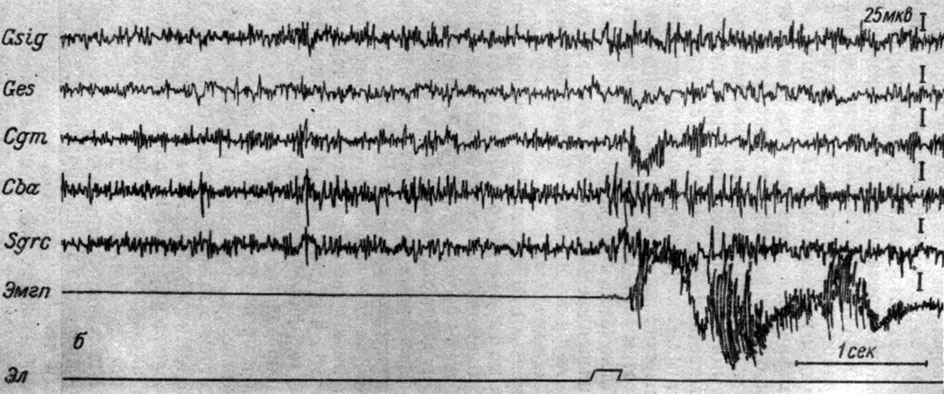
Рис. 43. Изменения ЭГ коры большого мозга и структур среднего мозга собаки № 9 (а) и собаки № 21 (б) при действии порогового электрического тока на правую переднюю лапу (а - опыт № 5 от 22/II 1958 г.; б - опыт № 12 от 12/IV 1958 г. - по Т. С. Наумовой, 1961)
Приведенные данные свидетельствуют о том, что электрические процессы в специфических и неспецифических (ретикулярных) системах каждого уровня мозга были, с одной стороны, очень сходными (т. е. широко иррадиированными) у каждого данного животного, а, с другой стороны, различными у разных животных. Так как наиболее широкий диапазон колебаний электрических потенциалов был обнаружен у наиболее подвижных животных (а двигательная активность собак, по данным М. С. Алексеевой и сотр., 1958, зависит от степени подвижности нервных процессов), можно думать, что у этих собак колебания глубины процессов возбуждения происходят в наиболее широких пределах.
Сходство изменений ЭГ в специфических и неспецифических системах ствола мозга у каждого данного животного позволяет заключить, что индивидуальные особенности нервной системы собак касаются всех мозговых структур и, видимо, создают соответствующую и одинаковую окраску процессов, определяющих характер эффекторных компонент реакции на раздражения (на то же указывают эксперименты Е. Ф. Мелиховой, 1953).
Генерализованные сдвиги электрических потенциалов у каждого животного имеют свои отличительные черты, и это дает основание полагать, что канву индивидуальных различий создает характер тех диффузных генерализованных процессов мозга, которые определяют целостную (со всеми ее компонентами) реакцию организма на раздражения.
Как хорошо известно, особенности формирования условных рефлексов существенно зависят от индивидуальных и типологических особенностей нервной системы животных. Так как смена стадий формирования рефлекса находит отражение в электрической картине активности корковых и стволово-подкорковых формаций, можно думать, что индивидуальные особенности высшей нервной деятельности выражаются в особенностях перераспределения процессов в этих формациях.
Как показали наши наблюдения, характер электрографической реакции, сопровождающей осуществление условного оборонительного рефлекса на каждом этапе его формирования, также зависел от индивидуальных особенностей нервной системы подопытных животных. Это выражалось в том, что у нескольких собак проявлялись не все электрографические фазы выработки условного оборонительного рефлекса (например, у 7 собак не была обнаружена I стадия, а у всех остальных животных они имели разную продолжительность (см. табл. V).
Кроме того, у разных животных обнаружена разная склонность к сохранению следов возбуждения, что проявляется в разной выраженности следовых и межсигнальных процессов как в кортикальных, так и субкортикальных структурах. Наиболее выраженные следовые реакции наблюдались у 7 собак, у других животных они были менее продолжительны и отчетливы.
Таким образом, индивидуальные особенности нервной системы собак определяются теми процессами, которые протекают как в коре головного мозга, так и в стволовых структурах. Индивидуальные различия существуют и в поведении параметров вызванных ответов в первичной проекционной зоне слуха не только до выработки оборонительных условных рефлексов, но также в процессе формирования последних. Так, например, начало ответа на первый щелчок у одного животного колебалось в пределах от 7-8 до 12 мсек, время до максимума позитивной волны - от 10 до 30 мсек и время до максимума негативной фазы - от 20 до 50-60 мсек. У другой собаки скрытый период первичного вызванного потенциала на первый щелчок испытывал колебания в пределах от 7-8 до 17 мсек. Время до максимума позитивной компоненты ответа колебалось в пределах 12-50 мсек и до максимума негативной волны - в пределах 25-60 мсек.
Особенно вариабельны были показатели амплитуды и длительности первых компонент вызванного ответа. Так, у одной собаки длительность первой позитивной компоненты вызванного потенциала менялась в пределах 6-40 мсек, длительность негативного отклонения колебалась от 12 до 60 мсек, и, наконец, длительность второй позитивной фазы находилась в пределах 12-72 мсек. У другой собаки длительность первой позитивной компоненты вызванного потенциала менялась в пределах от 30 до 80-90 мсек, длительность негативного отклонения колебалась от 18 до 60-70 мсек, длительность второй позитивной фазы находилась в пределах 6-40 мсек.
Индивидуальные особенности динамики вызванных ответов сказываются не только на различиях абсолютных значений их параметров, то также на последовательности этих изменений и их длительности в процессе выработки оборонительного условного рефлекса. Как следует, например, из рис. 19, у собаки № 3 почти не была выражена первая стадия формирования рефлекса; иными словами, образование последнего начиналось сразу с некоторого возрастания величины вызванных потенциалов в первичной проекционной слуховой зоне. У других животных (см. рис. 16, 22) в начале формирования условной связи, напротив, было отчетливо выражено резкое снижение амплитуды этих ответов. На тех же рисунках видно, как существенно различаются длительности каждой последующей электрографической стадии по этому показателю.
Приведенные факты говорят о существенных различиях в динамике вызванных потенциалов при становлении оборонительных условных рефлексов. Эти же факты дают основание полагать, что в известной мере противоречивость в утверждениях исследователей по обсуждаемому вопросу могла определяться индивидуальными особенностями нервной системы подопытных животных.
Следует особо подчеркнуть, что индивидуальные особенности динамики параметров вызванных потенциалов зависят не только от индивидуальных особенностей нервной системы животных, но также от положения отводящих электродов по отношению к фокусу максимальной активности.
Таким образом, генерализованные сдвиги электрических потенциалов у каждого животного имеют свои отличительные черты, что свидетельствует об индивидуальных различиях диффузных генерализованных процессов мозга, которые определяют индивидуальные особенности реакций основных и побочных компонент в целостной реакции организма на раздражения. Что же касается индивидуальных особенностей высшей нервной деятельности, то они выражаются в особенностях перераспределения электрической активности в анализаторных и межанализаторных структурах мозга, проявляющихся в длительности выделенных стадий формирования оборонительного условного рефлекса. Иными словами, приведенный материал с несомненностью свидетельствует об индивидуальных особенностях эффектов взаимодействия специфической и неспецифической афферентации на уровне специфических "заинтересованных" и "незаинтересованных" систем анализаторов и эффектов взаимодействия неспецифических афферентаций в каждой неспецифической формации мозга. У каждого данного животного обнаруживается одинаковая индивидуальная окраска специфической и неспецифической афферентации мозга, т. е. локальных и диффузных реакций, что дает основание думать об одинаковых индивидуальных особенностях нервных элементов структур всех уровней и систем мозга.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'