Глава IV. Взаимодействие специфической и неспецифической афферентации и его роль во второй период концентрации (IV стадии) условного оборонительного рефлекса и на V стадии формирования условного оборонительного рефлекса
Как широко известно, теория условных рефлексов И. П. Павлова предусматривает возможность разной структурной организации замыкательного процесса. Организация рефлекса, по-видимому, меняется по мере его выработки. Если на первых этапах становления рефлекса (в стадиях генерализации) условнорефлекторное возбуждение широко охватывает все субкортикально-стволовые и корковые формации мозга, а третья стадия (I период концентрации рефлекса) связана с участием систем взаимодействующих анализаторов, то по мере дальнейшего упрочения и известной автоматизации реакции наступает новое изменение организации условнорефлекторного возбуждения, впервые замеченное в опытах М. Н. Ливанова и К. Л. Полякова (1945), М. Н. Ливанова и сотр. (1951), В. В. Артемьева и Н. И. Безладновой (1952) и подтвержденное затем Morrell (1957), Gastaut и сотр. (1957), М. Я. Рабиновичем (1961) и многими другими исследователями. В этой стадии, обозначенной М. Н. Ливановым стадией концентрации оборонительного условного рефлекса, изоритмичная с условным (световым) раздражением электрическая активность появлялась только в сенсомоторной области коры мозга кролика. Аналогичное наблюдение было сделано В. В. Артемьевым и Н. И. Безладновой (1952), которые, исследуя электрические процессы в слуховой области коры мозга кошки при образовании условного оборонительного рефлекса на звук, обнаружили, что, когда количество сочетаний доходило до 250 и более, в отдельных случаях наблюдались условнорефлекторные реакции при отсутствии первичных электрических ответов слуховой проекционной области. В процессе выработки условного оборонительного движения происходит ограничение территории и изменение характера ответов, постепенно концентрирующихся в тех пунктах сенсомоторной зоны коры кошек, которые связаны с подкрепляющим движением (А. Б. Коган, 1960). Подтверждение данных, свидетельствующих об изменении локализации дуги рефлекса, получено и на собаках. Так, М. Я. Рабинович (1958, 1961) в стадии концентрации оборонительных условных рефлексов у собак наблюдал снижение уровня электрической активности в корковом конце анализатора, воспринимающего условный сигнал (слуховой коре). Г. Т. Сахиулиной было замечено в той же стадии выработки, и особенно при переключении, усиление активности лишь в лобно-теменной области мозга. В дальнейшем (Г. Т. Сахиулина, 1958) было уточнено, что активация наступала в двух участках: на границе gyr. coronalis и gyr, ectosylvius anterior и на стыке полей зрительного, слухового и соматосенсорного анализатора, в то время как в других зонах мозга электрические процессы изменялись мало. Тот же факт концентрации был обнаружен и на людях. Так, в исследованиях Gastaut и сотр. (1957), А. Гасто, А. Роже (1958) было обнаружено, что выработка речедвигательных условных рефлексов у человека сопровождается вслед за периодом генерализованных изменений ЭЭГ коры локальной блокадой аркообразного ритма сенсомоторной области коры, контралатеральной реагирующей конечности. С. М. Бабиян (1960) подтвердила эти наблюдения. В большой серии работ, посвященных исследованию частотно-специфического ответа на кроликах, кошках и обезьянах, также было показано, что при сочетании звука с ритмическим светом на смену генерализованной десинхронизации на звук приходят локальные процессы в зрительной зоне коры, которые сначала выражаются в воспроизведении звуком ритма светового раздражения, а затем локальной десинхронизации в этой области (Morrell, 1957; Morrell, Naguet, 1956; Morrell и сотр., 1957; Yoshii, Hockaday, 1958; Bergel и сотр., 1959; Morrell и сотр., 1960). Интересно отметить, что тот же порядок изменения ответов наблюдается и при сочетании индифферентного раздражения с ритмической электрической стимуляцией подкорковых структур. Так Yoshii и сотр. (1960) на неанестезированных кошках и собаках, используя раздражения ядерных структур (nucl. lateralis posterior, nucl. medialis dorsalis thalami, ретикулярной формации, миндалевидного ядра, nucl. caudatus, centrum medianum) в ритме 3-10 в 1 сек, которым предшествовали тон или свет, показали, что на смену генерализованным вызванным потенциалам с последующей генерализованной десинхронизацией коры приходят колебания в ритме 5 в 1 сек в соответствующих подкорковых структурах.
Таким образом, во всех этих исследованиях выступило то обстоятельство, что по мере упрочения реакции ответы на действие условного сигнала наблюдаются в корковом безусловнорефлекторном центре или центре, активирующемся при подкреплении, что в процессе упрочения оборонительного рефлекса "навязанные" ритмы наблюдаются только в сенсомоторной зоне коры и отсутствуют в зрительной области коры при действии ритмического светового условного раздражения. М. Н. Ливанов и сотр. (1951) считают, что путь условнорефлекторного возбуждения в процессе выработки вместо корково-коркового становится подкорково-корковым, ибо в наружном коленчатом теле продолжают регистрироваться ответы, следующие в ритме условной стимуляции. Исчезновение вызванных потенциалов из условнорефлекторного коркового очага при значительном упрочении условной реакции, по мнению В. В. Артемьева и Н. И. Безладновой (1952), также свидетельствует об изменении направления условнорефлекторного возбуждения, ибо появляющиеся в это время в сенсомоторной зоне коры вызванные ответы следуют с латентным периодом, более коротким, чем те, которые регистрируются в первичной проекционной зоне слуха. Д. С. Воронцов (1960) также разделял эту точку зрения, полагая, что многолетние следы не могут сохраняться в самих воспринимающих системах.
Приведенные выше электрофизиологические наблюдения позволили предположить, что в процессе выработки рефлекса происходит изменение его локализации. Если на первом этапе замыкание осуществляется по типу кора-кора, в дальнейшем же оно происходит, видимо, без активного участия коркового уровня сигнального анализатора. Отсутствие условнорефлекторных изменений активности в кортикальных звеньях сигнального анализатора во второй стадии концентрации свидетельствует о том, что эти звенья прекращают играть активную роль в формировании временной связи.
Встает, естественно, вопрос о том, какими путями в последнем случае распространяется условнорефлекторное возбуждение с субкортикального уровня сигнального анализатора к корковому безусловнорефлекторному центру. Нужно признать, что путей проведения от стволового уровня зрительного, слухового и других анализаторов к сенсомоторной коре достаточно много. Но в связи с тем, что некоторые из ядер таламической диффузной проекционной системы локально проецируются в сенсомоторную зону коры, А. Гасто и А. Роже (1958) полагают, что условнорефлекторное возбуждение к коре следует по неспецифическим путям, ранее участвовавшим в передаче безусловного сигнала. Последнюю точку зрения разделяют Yoshii и сотр. (1958а), Н. Иошии и сотр. (1958б), которые пришли к выводу, что при образовании оборонительных и пищевых условных рефлексов у собак, а также условнорефлекторном вызове так называемых частотно-специфических ответов, замыкание осуществляется благодаря тому, что условнорефлекторное возбуждение достигает безусловного коркового центра с помощью таламической неспецифической проекционной системы. В отношении пищевых условных рефлексов определенно можно думать, что условнорефлекторное формирование активности коркового пищевого центра, расположенного в gyr. orbitalis и gyr. sylvius anterior, идет с участием некоторых неспецифических ядер зрительного бугра (Л. Г. Трофимов и Н. Н. Любимов, 1960).
Прямых доказательств участия ретикулярных структур мозга и других межанализаторных систем в проведении возбуждения из субкортикальных отделов анализатора условного сигнала в кортикальные зоны кожно-двигательного анализатора на стадии упроченных оборонительных рефлексов пока нет, но такое предположение имеет основания.
Как показали наши наблюдения, активация процессов на действие условного сигнала в системе сигнального анализатора по мере сочетаний становится все менее и менее выраженной. В системе слухового анализатора нарастание возбуждения почти не проявляется, и изменения электрической активности в кохлеарных ядрах, внутреннем коленчатом теле, коре эктосильвиевой и сильвиевой извилин становятся аналогичными сдвигам, происходящим в других структурах мозга.
На рис. 15, а, где представлена динамика амплитудной характеристики электрографической реакции разных формаций мозга при образовании условного оборонительного рефлекса на тон 500 гц 48 дб у собаки № 4, видно, что в IV стадии формирования условного рефлекса происходит постепенное уменьшение степени усиления процессов возбуждения в системе слухового анализатора и, в частности, в его первичных переключательных реле - кохлеарных ядрах. Нивелировка изменений ЭГ в кохлеарных ядрах делает сходными процессы в кохлеарных ядрах, ядрах нежного и клиновидного канатиков, нижнем центральном ядре ретикулярной формации, которые выражаются в незначительном возрастании активности. Несколько снижается и уровень активирования коркового конца кожно-двигательного анализатора (рис. 15, б).
В IV стадии образования условного оборонительного рефлекса включение тона 500 гц 48 дб у собаки № 4 (см. рис. 16) сопровождается появлением вызванных потенциалов в коре средней эктосильвиевой извилины, которые по величине значительно уступают ответам в предыдущий период формирования рефлекса.
Что касается следовых процессов, то на IV стадии выработки рефлекса у той же собаки они также становятся значительно менее выраженными как в стволовых (см. рис. 17, а), так и в кортикальных структурах (см. рис. 17, б). Их длительность в тех и других образованиях не превышает 1 сек (см. рис. 18).
Сопоставление аналогичных изменений электрической активности разных структур мозга собаки № 3 с вызванными ответами, появляющимися на включение условного сигнала (тона 500 гц 48 дб) в ЭЭГ первичной проекционной корковой зоны слуха, показало, что в IV фазе выработки рефлекса происходит постепенное снижение амплитуды вызванных потенциалов. Сказанное иллюстрирует рис. 19, на котором видно, что в IV стадии образования условного рефлекса уменьшаются вызванные ответы не только в коре эктосильвиевой извилины, но также в коре сигмовидной извилины по сравнению с ответами в III фазе выработки рефлекса.
Снижение уровня активации бульбарных и кортикальных звеньев сигнального анализатора по мере упрочения условного рефлекса может выражаться и в появлении медленных колебаний, однако последние охватывают не все его звенья. На уровне продолговатого мозга в трапециевидном теле - верхней оливе, - медленные волны не регистрируются, в них происходит усиление процессов возбуждения. Экзальтация активности наступает и в крупноклеточном ядре ретикулярной формации.
Количественный анализ изменений электрической активности, наступающих при действии условного сигнала в IV стадии его образования у собаки № 11, показал, что в этот период уменьшается степень активации кохлеарных ядер (рис. 30, 38) и коры сильвиевой извилины, ЭЭГ которой отводится с помощью погружных электродов. Более того, в этих формациях в ответ на действие условного сигнала появляются медленные волны (показаны на графиках изменений уровня активности кохлеарных ядер кружками). Одновременно в крупноклеточном ядре ретикулярной формации регистрируется эффект активации. Следовые процессы в исследованных образованиях на этом этапе формирования временной связи электрографически не проявляются.
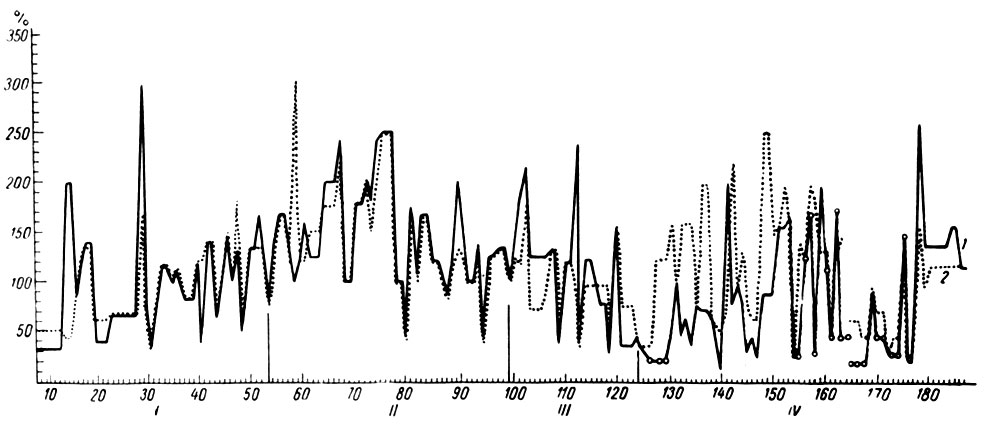
Рис. 38. Амплитудная характеристика ЭГ условной реакции при формировании оборонительного рефлекса на тон 500 гц 100 дб (собака № 11). Обозначения те же, что на рис. 30
Особенно отчетливо появление медленных колебаний в системе сигнального анализатора и эффект активации в гигантоклеточном ядре ретикулярной формации обнаружились у той же собаки № 11 при формировании условного рефлекса на более сильные звуковые сигналы (тон 500 гц 100 дб) (см. рис. 38). В третьей стадии выработки этого рефлекса условная реакция протекала на фоне слабой экзальтации активности (рис. 39, а, 124-е сочетание), которая следовала за кратковременным (250 мсек) периодом генерализованной депрессии. Нарастание осцилляций в кохлеарных ядрах и слуховой зоне коры было несколько более выражено, чем в других территориях. Уже при 125-м сочетании (рис. 39, б) кратковременный период генерализованного угнетения электрических колебаний сопровождается их усилением в коре коронарной извилины, крупноклеточном ядре ретикулярной формации и трапециевидном теле на границе с верхнеоливарным комплексом ядер. Напротив, в кохлеарных ядрах и коре сильвиевой и эктосильвиевой извилин наблюдается появление медленных волн в ритме 5-6 в сек, перемежающихся с периодами депрессии активности. Поражает удивительное сходство в процессах, происходящих в крайних точках слухового анализатора (первичном переключательном реле и корковом его конце).
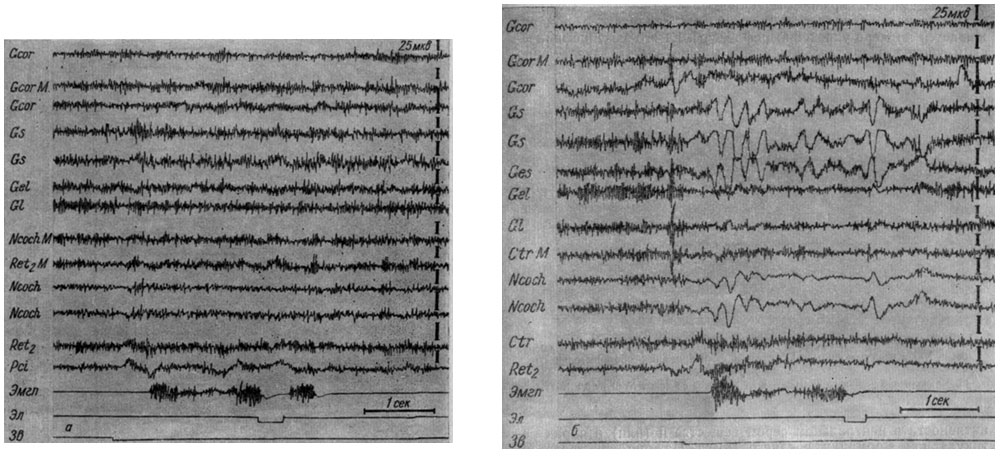
Рис. 39. Изменения ЭГ структур коры и продолговатого мозга собаки № 11 наступающие в IV стадии образования условного оборонительного рефлекса на тон
Дезактивация процессов в слуховом анализаторе обнаруживается по мере упрочения рефлекса и на его мезенцефалическом уровне. Параллельно с этим можно видеть экзальтацию активности в центральном сером веществе и (или) в ядрах ретикулярной формации среднего мозга. На рис. 40 представлены ЭГ структур среднего мозга и коры собаки № 2 при 166-м применении условного сигнала (тона 500 гц 48 дб). Видно, что через 0,3 сек после начала действия звука наступает снижение величины электрических осцилляций в коре супрасильвиевой, коронарной извилин, внутреннем коленчатом теле (в последнем обнаруживаются длительные смещения уровня активности) и дорсальном отделе ретикулярной формации среднего мозга (nucl. cuneiformis). Периоду депрессии активности в указанных структурах соответствует экзальтация ее в центральном сером веществе среднего мозга и эффект прерывистой активации на уровне заднего двухолмия. Отмеченные изменения предшествуют появлению выраженной моторной реакции реагирующей конечности. В последействии рефлекса происходит процесс генерализованного угнетения электрических потенциалов, и лишь в записях ЭЭГ коры коронарной извилины и ЭГ центрального серого вещества ствола сохраняются более высокоамплитудные, чем в других зонах мозга, колебания. Описанные изменения резко отличаются от наблюдавшихся на III стадии образования рефлекса (см. рис. 32).
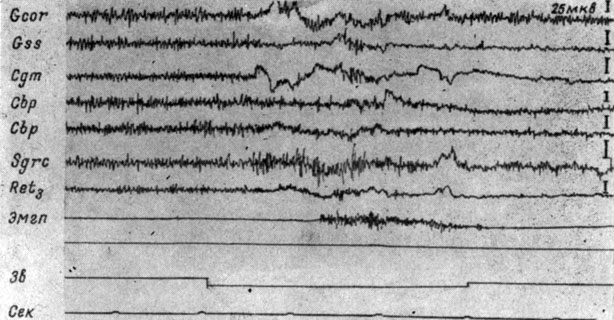
Рис. 40. Изменения ЭГ структур коры большого и среднего мозга собаки № 2, наступающие в IV стадии формирования оборонительного условного рефлекса. 166-е применение условного сигнала - тона 500 гц 48 дб (опыт № 61 от 3/I 1959 г.)
На рис. 41 иллюстрируются изменения ЭГ на уровне среднего мозга и коры при осуществлении условного рефлекса на IV стадии его выработки у собаки № 16. Видно, что при 174-м применении условного сигнала (тон 500 гц 48 дб) вместо преимущественного усиления электрических потенциалов в коленчатом теле, которое имело место в III стадии образования рефлекса, и, в частности, при 62-м применении условного раздражения (рис. 33, б), наступает экзальтация электрических процессов в ретикулярной формации (nucl. cuneiformis) и центральном сером веществе среднего мозга. В сенсомоторной и слуховой зонах коры при этом происходит реакция десинхронизации (снижение амплитуд и нарастание ритма электрических осцилляций), а во внутреннем коленчатом теле, заднем двухолмии и красном ядре - эффект нарастания ритма колебаний.
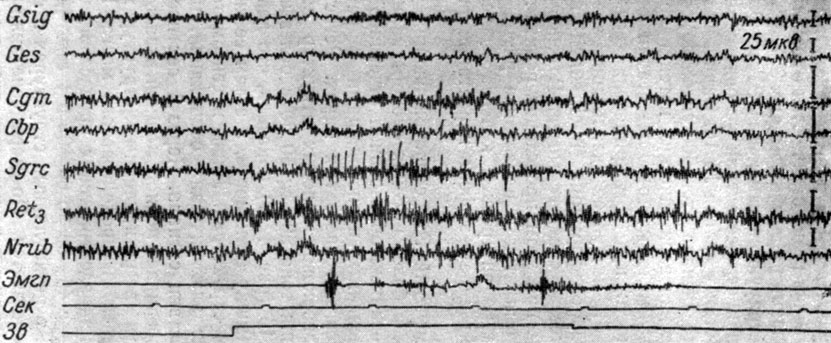
Рис. 41. Изменения ЭГ структур коры большого мозга и среднего мозга собаки № 16, наступающие в IV стадии формирования оборонительного условного рефлекса. 174-е применение условного сигнала - тона 500 гц 48 дб (опыт № 69 от 20/VI 1958 г. - по Т. С. Наумовой, 1965)
Что касается изменений ЭГ, сопровождающих IV стадию выработки рефлекса, в более высоких субкортикальных структурах, то из обследованных образований отчетливо выявилась связь между изменениями, происходящими в ретикулярной формации ствола мозга и хвостатых ядрах. Регистрация ЭГ коры и разных подкорково-стволовых структур во время и после 133-го сочетания тона 500 гц 30 дб с болевым подкреплением у собаки № 15 (рис. 42, а) показала, что действие тона вызвало сначала вспышку активности, а затем ее снижение во всех регистрируемых структурах. Лишь в сенсомоторной зоне коры наступает рост электрических осцилляций, продолжающийся и в последействии сигнала. Через 1 сек после прекращения стимуляции возникает период мощной экзальтации электрической активности в ретикулярных структурах среднего (где он особенно силен) и продолговатого мозга. По сравнению с фоном величина электрических процессов растет в 4-5 раз, а их частота - с 45-50 до 60-65 в 1 сек. Через 2,5 сек активация процессов в ретикулярных структурах и сенсомоторной зоне коры прекращается. По мере сочетаний описанные следовые процессы начинают регистрироваться в ответ на действие самого сигнала. На рис. 42, б представлено 171-е сочетание тех же раздражителей у той же собаки. Отчетливо видно, что звук вызывает слабую десинхронизацию активности в корковых структурах, а в хвостатом ядре - возрастание ритма (с 48-50 до 55-58 в 1 сек) и величины (почти вдвое) электрических колебаний, которое продолжается и после осуществления рефлекса. В ретикулярной формации среднего мозга, (nucl. subcuneiformis), а также черной субстанции частота колебаний увеличивается от 45-50 до 58-60 в 1 сек, а их амплитуды достигают 70-80 мкв (при величине их в фоновой записи 30-35 мкв). В прочих образованиях отмечается эффект слабого возбуждения. Через 5 сек в последействии сочетания регистрируется остаточная активация преимущественно в коре, хвостатых ядрах, ретикулярной формации и пирамидах.
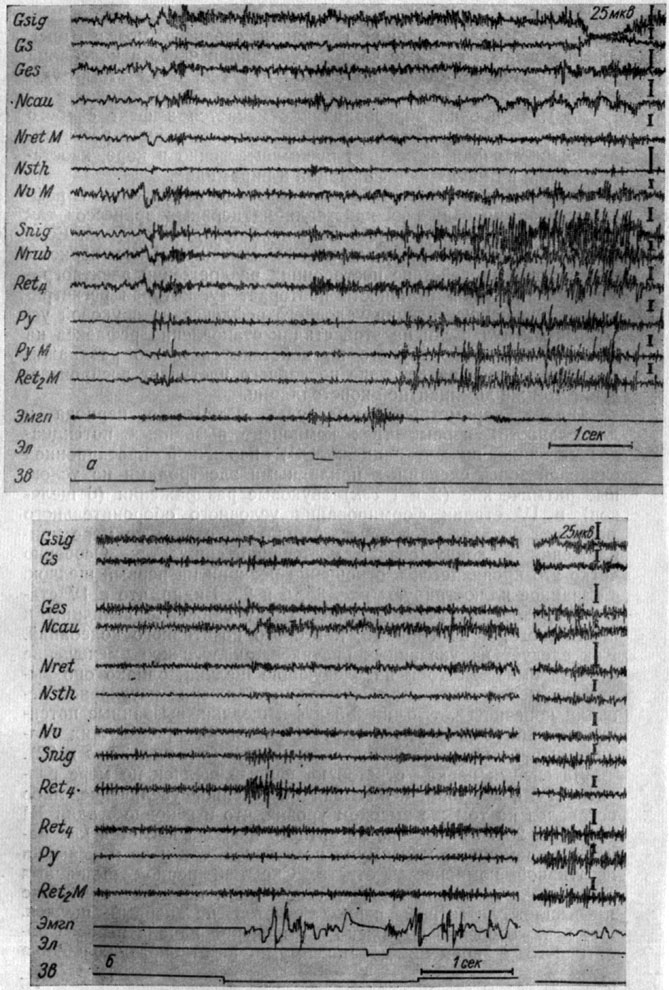
Рис. 42. Изменения ЭГ коры большого мозга и подкорково-стволовых структур собаки № 15, наступающие в IV стадии формирования оборонительного условного рефлекса. а - 133-е, б - 171-е сочетание тона 500 гц 30 дб с электрокожным раздражением правой передней лапы (а - опыт № 44 от 2/I 1962 г.); б - опыт № 64 от 23/II 1962 г - по Т. С. Наумовой, 1965
В IV стадии выработки оборонительного условного рефлекса сформировавшаяся констелляция нервных процессов так же, как и в предыдущих стадиях, является доминантной. Если движение конечности происходит в межсигнальные периоды или в ответ на действие посторонних раздражений электрографическая картина активности повторяет ту, которая регистрируется при осуществлении условного рефлекса на звуковой условный стимул. Если на этой стадии становления рефлекса на звук начать выработку его на действие светового раздражителя, то в ответ на действие последнего наступает воспроизведение той же доминантной констелляции.
Анализ параметров (амплитуды, латентного периода и длительности) первых трех компонент вызванных потенциалов, возникающих в записях ЭЭГ первичной проекционной слуховой зоны, сделанных накладными электродами на условные ритмические (2 в 1 сек) звуковые раздражения (6 щелчков) в IV стадии формирования условного оборонительного рефлекса показал следующее. Амплитуда ответа по мере приближения к концу стадии постепенно снижается. Особенно четко это проявляется в отношении реакции на первый щелчок. Сказанное иллюстрируют рис. 35, б и 22. Видно, что в IV стадии образования рефлекса происходит снижение величины и позитивного, и негативного, и второго позитивного колебаний. Как следует из указанных рисунков, аналогичное уменьшение ответов имеет место и на следующие щелчки, однако оно выражено менее отчетливо. По мере приближения к условной реакции конечности, т. е. на 3-й и 4-й щелчки, вызванные потенциалы либо также уменьшаются, как и на предыдущих стадиях выработки, либо иногда возрастают (см. рис. 37, в и 22).
Латентный период ответов на первый щелчок по мере развития IV фазы либо испытывал постепенное увеличение, либо оставался на том же высоком уровне, что и в конце стадии III (см. рис. 23). Увеличение латентного периода касается как начала и времени до максимума позитивной, так и до максимума негативной компоненты ответа. Скрытый период вызванных ответов на третий и четвертый щелчки к концу стадии даже превышает латентный период ответов на первый щелчок. Это было особенно выражено в отношении начала первого позитивного и времени до максимума негативного колебания вызванного ответа у собаки № 20 (см. рис. 36). На этом графике видно, что начиная с 155-го сочетания двигательная условная реакция сопровождается возрастанием латентного периода вызванных потенциалов.
Одновременно с удлинением латентного периода и уменьшением величины вызванного ответа на первый щелчок из 6, служащих сигналом условной реакции в IV фазе формирования рефлекса, происходит дальнейшее увеличение его длительности. Увеличение длительности касается всех компонент вызванного потенциала (см. рис. 24). Длительность фаз вызванного ответа, так же как и латентный период и амплитуда, меняется и в момент, предшествующий и сопутствующий условнорефлекторной реакции конечности, а именно, если в первых трех стадиях формирования рефлекса условнорефлекторная реакция сопровождалась сокращением, то в IV стадии образования условного рефлекса нередко можно наблюдать увеличение длительности всех компонент вызванного ответа. На рис. 37, в видно, что при 162-м сочетании 6 щелчков и болевого подкрепления амплитуда и длительность компонент вызванного потенциала на 5-й щелчок, совпадающий с условной реакцией, не уменьшается, как в предыдущей стадии (см. рис. 37, б), а, наоборот, увеличивается. Описанные изменения длительности вызванных потенциалов предшествуют их исчезновению.
Итак, изменения ЭГ, сопровождающие IV стадию выработки оборонительного условного рефлекса на звуковые сигналы, выражаются в появлении процессов, охватывающих несколько иную, чем в III стадии, констелляцию нервных центров. Условная реакция на звуковой сигнал подпороговой, пороговой и надпороговой интенсивности на IV этапе выработки рефлекса формируется на фоне локальной активации (возрастания ритма и амплитуд колебаний) электрических процессов, обнаруженной в записях ЭГ погружными электродами от комплекса трапециевидное тело - верхняя олива, крупноклеточного ядра ретикулярной формации продолговатого мозга, ряда ядер ретикулярной формации среднего мозга (nucl. cuneiformis, nucl. subcuneiformis), черной субстанции, центрального серого вещества и хвостатых ядер. Изменения ЭЭГ сенсомоторной зоны коры, наступающие в ответ на действие условного звукового сигнала в записях накладными электродами, выражаются в десинхронизации активности (снижении амплитуд и возрастании ритма электрических колебаний). Лишь у двух животных регистрировался эффект экзальтации электрических осцилляций.
Активация процессов в новой констелляции структур появляется сначала на следах условных рефлексов и лишь затем обнаруживается и в ответ на действие условного сигнала, а также и в межсигнальные периоды.
Новая констелляция, которая создается на IV стадии образования рефлекса, имеет доминантные свойства - она воспроизводится в начале формирования условного оборонительного рефлекса на ритмический свет.
Параллельно с нарастанием уровня возбуждения в трапециевидном теле - верхней оливе, ретикулярной формации (nucl. gigantocellularis, nucl. cuneiformis, nucl. subcuneiformis), черной субстанции, центральном сером веществе, хвостатых ядрах, сенсомоторной зоне коры в момент осуществления условного рефлекса на звук, а также в межсигнальные периоды отсутствуют существенные изменения активности как в большинстве ядер слухового анализатора (кохлеарных ядрах, внутреннем коленчатом теле, коре эктосильвиевой и сильвиевой извилин), так и в других территориях мозга, как-то: гиппокампе, ядрах таламуса, ножках мозжечка, ядрах тройничного нерва и т. п. Осуществление условного рефлекса может сопровождаться появлением медленных волн в системе слухового анализатора (кохлеарных ядрах, внутреннем коленчатом теле, коре сильвиевой, эктосильвиевой извилин).
Локальные процессы наиболее четко выявляются при биполярном способе записи ЭГ погружными электродами. Исчезнувшую локальную активацию ядер слуховой системы на действие условного сигнала можно, однако, вновь вызвать, если начать выработку рефлекса на звуковое раздражение другой частоты или интенсивности. Анализ амплитудной характеристики первых трех компонент вызванного потенциала на условные ритмические (2 в 1 сек) звуковые раздражения (щелчки) в IV стадии формирования рефлекса показал, что амплитуда позитивной, негативной и второй позитивной компоненты ответа по мере приближения к концу стадии постепенно снижается. Особенно четко это проявляется в отношении ответа на первый щелчок.
Латентный период начала и время до максимума позитивной и максимума негативной компоненты ответа либо остается на том же высоком уровне, что и в конце III стадии, либо испытывает постепенное увеличение. Увеличение латентного периода касается начала и времени до максимума позитивной, так и до максимума негативной компоненты ответа и также особенно четко выражено в отношении реакции на первый щелчок.
Одновременно с удлинением латентного периода и уменьшением величины вызванного ответа на первый щелчок в IV фазе формирования рефлекса происходит дальнейшее увеличение его длительности. Увеличение длительности касается всех компонент вызванного потенциала, оно особенно выражено к концу IV периода образования рефлекса.
Амплитуда ответов, латентный период их появления, а также длительность меняются и в момент, предшествующий или сопутствующий началу условной оборонительной реакции (т. е. например, на третий, четвертый щелчки). Амплитуды первых трех компонент вызванных ответов перед двигательной реакцией либо уменьшаются, так же как на предыдущих стадиях образования рефлекса, либо (в некоторых случаях) возрастают. Латентный период начала, время до максимума первой позитивной и до максимума негативной компоненты, а также длительность первых трех компонент ответа на третий-четвертый щелчки, которые следуют перед осуществлением двигательной условной реакции, превышают скрытый период появления и длительность тех же компонент вызванного потенциала на первый щелчок.
Четвертая стадия образования рефлекса совпадала с периодом стандартизации латентного периода эффекторной реакции и периодом наиболее прочных условнорефлекторных ответов.
Как показывают наши данные, на IV стадии выработки рефлекса, т. е. по мере известной автоматизации, так же как в опытах М. Н. Ливанова и сотр. (1951), В. В. Артемьева и Н. И. Безладновой (1952), М. Я. Рабиновича (1961) и других, наступает постепенная нивелировка процессов в корковом конце сигнального (слухового) анализатора, которая этими авторами расценивается как следствие отклонения пути условнорефлекторного возбуждения и развития процесса торможения в этом анализаторе.
По нашим наблюдениям, уменьшение вызванных потенциалов происходит при одновременном удлинении латентного периода и длительности вызванных ответов на первые щелчки в серии, служащей сигналом условной оборонительной реакции (см. рис. 28). Эти данные говорят о том, что механизмы исчезновения вызванных ответов в момент эффекторной реакции в первых трех стадиях выработки рефлекса и механизмы исчезновения вызванных ответов на включение условного сигнала в IV стадии его формирования совершенно различны. Если в первом случае речь шла о маскировке вызванных ответов, то во втором случае исчезновение вызванных потенциалов связано с постепенным понижением возбудимости сигнального анализатора к действию условного раздражения.
Понижение возбудимости сигнального анализатора проявляется также в процессе действия условного сигнала и особенно в момент осуществления эффекторной реакции на той же стадии формирования рефлекса, в момент, непосредственно предшествующий и сопровождающий движение конечности, нередко происходит удлинение латентного периода и длительности вызванных потенциалов в ответ на 3, 4-й щелчки по сравнению с параметрами ответов на первые щелчки в серии из 6 щелчков, служащих сигналом условной реакции (см. рис. 28). Понижение возбудимости сигнального анализатора в IV стадии образования рефлекса свидетельствует о постепенном развитии эффекта привыкания к действию условных сигналов.
По мере значительного упрочения реакции эффект активации исчезает не только из коркового уровня слухового анализатора, но также из внутренних коленчатых тел, заднего двухолмия и кохлеарных ядер. Более того, иногда удается видеть почти во всей слуховой системе медленные волны, свидетельствующие о развитии здесь тормозного процесса в то время, как в верхнеоливарном комплексе ядер, трапециевидном теле наступает отчетливый эффект активации (эффект нарастания возбуждения). Эти наблюдения свидетельствуют о том, что основной поток информации, формирующий доминантную констелляцию в этом случае, осуществляется, по-видимому, через трапециевидное тело и верхний оливарный комплекс ядер. Значение верхней оливы как одного из важных участников условнорефлекторного процесса подтверждается и данными Worden (1959), который показал, что величина вызванных первичных ответов в слуховой системе (начиная от кохлеарных ядер, трапециевидного тела и кончая корой) зависит от периода выработки условной реакции (в ответ на звуковые тоновые ритмические сигналы - нажатие на педаль, подкрепляемое пищей). На первых этапах ответы наблюдались сначала главным образом в верхней оливе, потом возникали генерализованные по всей слуховой системе. По мере упрочения условной реакции на звук вызванные потенциалы постепенно исчезали из всех ядер слухового анализатора, оставаясь лишь в задних отделах верхнеоливарного комплекса. Усиление процессов в верхнеоливарном комплексе ядер при снижении возбуждения в кохлеарных ядрах как будто свидетельствует о возможном ответвлении пути условнорефлекторного возбуждения на уровне верхних олив. Каким образом в этом случае возбуждение не отражается в ЭГ кохлеарных ядер - остается загадкой.
М. Н. Ливанов и сотр. (1951) считали, что перемещение дуги условного рефлекса и превращение из корково-коркового типа замыкания в подкорково-корковый происходит на уровне межуточного мозга. Ими было замечено, что в стадии концентрации рефлекса вызванные световой стимуляцией потенциалы перестают регистрироваться в зрительной зоне коры мозга кролика, но продолжают наблюдаться в наружном коленчатом теле и сенсомоторной зоне коры головного мозга. Нам не удалось найти коррелятивной связи между процессами во внутреннем коленчатом теле и сенсомоторной зоне коры головного мозга в IV стадии выработки условного оборонительного рефлекса у собак. В этом подкорковом звене слухового анализатора регистрировались те же процессы дезактивации, что и в его корковом конце. Что касается заднего двухолмия, которому ряд авторов придает большое значение в условнорефлекторной деятельности (Kryter, Ades, 1943; Т. А. Меринг, 1960, и др.), то в нем у некоторых животных и в отдельные периоды четвертой стадии выработки рефлекса удавалось видеть больший уровень активации, чем во внутреннем коленчатом теле и слуховой зоны коры, однако эти наблюдения требуют подтверждения и дальнейшего изучения. Не исключена возможность, что существует известная временная последовательность включения того и другого уровня слухового анализатора в реакцию. Можно думать, что сначала активным участником условнорефлекторного процесса является заднее двухолмие и лишь затем верхний оливарный комплекс. Однако это предположение нужно рассматривать лишь как сугубо предварительное.
На какие же образования может переключаться условнорефлекторное возбуждение с нижних звеньев сигнального анализатора? По-видимому, в первую очередь на сенсомоторную зону коры.
Изменения ЭЭГ сенсомоторной зоны коры, наступающие в ответ на действие условного звукового сигнала в IV стадии, выражаются либо в эффекте десинхронизации активности (большинство животных), либо в эффекте экзальтации электрических осцилляций (два животных). Возможны два предположения по поводу этих различий в реакциях сенсомоторной зоны коры у разных животных в стадии значительно упроченных рефлексов. Эти различия либо связаны с тем, что у животных с выраженной экзальтацией электрических колебаний в сенсомоторной зоне коры в ответ на действие условного сигнала еще не достигнута достаточная степень упрочения рефлекса, либо что различия в реакциях связаны с разной локализацией отводящих электродов. Оба предположения кажутся вероятными. Наши исследования показали, что на IV стадии выработки рефлекса условнорефлекторная реакция, кроме сдвигов ЭЭГ ряда звеньев слухового анализатора и сенсомоторной зоны коры, начинает сопровождаться преимущественными изменениями электрической активности в ретикулярных структурах ствола мозга и хвостатых ядрах, в которых регистрируется значительный эффект активации, в то время как в других системах не выявляется никаких видимых изменений потенциалов. Активация ретикулярных структур происходит на уровне гигантоклеточного ядра продолговатого мозга, на уровне центрального серого вещества и ядер среднего мозга (nucl. cuneiformis, nucl. subcuneiformis).
Полученные до сих пор данные (см. обзор Т. С. Наумовой, 1963) показали отсутствие существенной разницы в состоянии условных рефлексов у интактных животных и животных с частично разрушенной ретикулярной формацией. То же следует и из экспериментов с их раздражением. Стимуляция ретикулярной формации вызывает лишь некоторое облегчение и ускорение или торможение условных реакций, т. е. влияния, которые, по-видимому, имеют неспецифический характер. Эти наблюдения свидетельствуют о том, что участие ретикулярных структур в замыкании, очевидно, не является определяющим. Но, вместе с тем, неспецифические системы не могут не принимать участия в осуществлении условного рефлекса. Так как ретикулярная формация является субстратом осуществления рефлекторных (вегетативных и соматических) функций нервной системы, а в связи с этим и субстратом реализации (а возможно и замыкания) побочных компонент условных рефлексов (П. К. Анохин, 1958, 1962; Л. Г. Трофимов и сотр., 1958), она является главным (но, как уже говорилось, не единственным) субстратом проведения неспецифической афферентации коры большого мозга, которая облегчает взаимодействие анализаторов, и, наконец, ретикулярная формация является одним из важных звеньев в осуществлении эффекторных функций мозга.
С другой стороны, с ее участием проводится болевая афферентация, а именно ее проведение осуществляется спиноталамическим трактом, ретикулярной формацией ствола (С. Дж. Херрик, Г. Г. Бишоп, 1958; В. Г. Агафонов, 1960), центральным серым веществом (Melzack и сотр., 1958), парафасцикулярным комплексом ядер таламуса (В. Г. Агафонов, 1960). Болевая афферентация опосредуется адренергическими элементами (И. П. Анохина, 1956; В. Г. Агафонов, 1956; П. К. Анохин, 1957, и др.). То обстоятельство, что проведение болевой чувствительности осуществляется с участием путей и ядер, расположенных в ретикулярной формации ствола мозга, определяет особую специфичность участия этой формации в осуществлении оборонительных условных рефлексов. Структуры ретикулярной формации, которые причастны к проведению болевой афферентации, в отношении оборонительного условного рефлекса представляют подкорково-стволовое звено безусловнорефлекторного анализатора. В таком смысле эти структуры входят в состав кожно-двигательного анализатора. Чем же объяснить, что при разрушениях ретикулярной формации не происходит исчезновения условных оборонительных рефлексов. По-видимому, это происходит вследствие того, что протопатическая афферентация коры осуществляется не только ретикулярной формацией, но по аналогии с диффузной афферентацией - самыми различными системами мозга. Это заключение делается на основании тех данных и соображений, которые были приведены в предыдущих главах.
Наблюдение преимущественной активации ретикулярной формации в IV стадии выработки рефлекса могло бы быть расценено как факт, подтверждающий мнение тех исследователей (Yoshii и сотр., 1956; Hernandez-Peon, 1957; Gastaut, 1955; Г. Гасто, 1958; Fessard, Gastaut, 1958, и др.), которые придают главное значение ретикулярной формации в замыкании. Так ли это? Приведенный нами материал с несомненностью свидетельствует, что формирование временной связи происходит вследствие взаимодействия анализаторов на уровне каждого их звена, где, таким образом, осуществляется встреча возбуждений, вызванных с сигнального и безусловнорефлекторного анализаторов. В этом смысле те процессы, которые разыгрываются в ретикулярной формации, могут быть расценены как эффект взаимодействия возбуждения слухового и болевого происхождения на уровне подкорковых звеньев кожно-двигательного анализатора. Это взаимодействие осуществляется одновременно в хвостатых ядрах и сенсомоторной зоне коры головного мозга. Активация процессов в системе ядер ретикулярной формации не может быть расценена как доказательство определяющей роли процессов конвергенции на уровне этой формации, так как все имеющиеся данные (см. предыдущий раздел) свидетельствуют о том, что кортикальные компоненты кожно-двигательного анализатора являются главным субстратом взаимодействия анализаторов при формировании оборонительного условного рефлекса.
Пока неясно, за счет процессов в каких компонентах (афферентных или эфферентных) происходит активация, регистрируемая на уровне ретикулярной формации в IV стадии выработки оборонительного условного рефлекса. Для ответа на этот вопрос требуются специальные исследования. Но в связи с тем, что ретикулярная формация представляет субстрат рефлекторных реакций, а также аппарат, трансформирующий тонизирующие и координирующие корковые и субкортикальные влияния на мышечную деятельность, вегетативные функции организма и восприятие, трудно исключить мысль, что нарастание уровня возбуждения в системе ретикулярная формация - хвостатое ядро в период осуществления условного оборонительного рефлекса в IV стадии его формирования связано с активацией эфферентных элементов. Однако появление стойкого очага возбуждения на границе лобной и теменной областей при упрочении и переключении условных оборонительных рефлексов, наблюдавшееся в опытах Г. Т. Сахиулиной (1957) (что, очевидно, является отражением усиления неспецифической афферентации в этой стадии выработки рефлекса), свидетельствует о том, что активация в системе ретикулярных структур имеет скорее афферентную природу.
Слуховое возбуждение проводится к системе кожно-двигательного анализатора различными путями. Как известно, слуховой анализатор на уровне ствола мозга связан многочисленными связями с самыми различными и в первую очередь ретикулярными ядрами.
Что касается проведения ретикулярного возбуждения в кору головного мозга, то на основании морфологических данных и физиологических наблюдений разные авторы предполагают 6 возможных путей (см. обзор Т. С. Наумовой, 1963). Известно, что в ретикулярной формации имеются клетки с длинными восходящими аксонами, доходящими до неспецифических таламических ядер, гипоталамуса, субталамуса, хвостатых ядер, структур обонятельного мозга и так далее, через которые, видимо, и осуществляются ретикулярные влияния на кору мозга. Кроме того, предполагается, что имеются и прямые пути, идущие от ретикулярной формации в кору головного мозга (М. Е. Шайбель, А. Б. Шайбель, 1958).
Несмотря на обилие связей ретикулярной формации с корой, в IV стадии выработки рефлекса используются, по-видимому, далеко не все. Исследование электрической активности структур межуточного мозга и подкорковых образований обнаружило, что в четвертой стадии выработки рефлекса параллельно с нарастанием возбуждения в ретикулярной формации ствола мозга происходит нарастание его в хвостатом ядре. По-видимому, ретикуло-стриарно-корковые пути, показанные в отношении ретикуло-стриарных путей М. Е. Шайбель, А. Б. Шайбель (1958) и в отношении стрио-кортикальных связей Monakow (1895), играют определенную роль в замыкании оборонительных условных рефлексов в стадии их значительного упрочения. Эксперименты с разрушением хвостатых ядер показали, что это вмешательство ведет к полному расстройству высшей нервной деятельности (Б. Н. Клосовский, Н. С. Волжина, 1956; Dean, Davis, 1959) или временным ее изменениям (Rosvold, Delgado, 1956; У. Р. Инграм, 1958). Торможением оборонительных и пищедобывательных безусловных и условных реакций сопровождается и раздражение этих ядер (Н. И. Лагутина, 1949, 1955; Н. А. Рожанский и Н. И. Лагутина, 1957; Nakao, Maki, 1958; В. А. Черкес, 1958; Umbuch, 1959). Высокочастотная стимуляция хвостатых ядер вызывает выраженный эффект коркового возбуждения - десинхронизацию ЭЭГ (Shimamoto, Verzeano, 1954; Stoupel, Terzuolo, 1954; Buchwald, Wyers, 1959). Если раздражение хвостатых ядер проводится редкими стимулами, в сигмовидной извилине коры наступает явление вовлечения (Buchwald, Wyers, 1959), которое рассматривается как проявление тормозного влияния. Таким образом, хвостатые ядра обладают способностью и тормозить и возбуждать деятельность коры (Monnier и сотр., 1959). На основании этих данных Shimamoto и Verzeano (1954), Б. Н. Клосовский и Н. С. Волжина (1956) полагают, что функция хвостатых ядер сводится к посылке дополнительных импульсов, нормализующих процессы возбуждения и торможения в коре больших полушарий, и в этом отношении функциональное значение хвостатых ядер и неспецифических систем ствола сходно.
Хотя нет физиологических доказательств специфической причастности хвостатого ядра к проведению болевой чувствительности, однако есть морфологические данные, свидетельствующие о том, что спино-таламический тракт, проводящий болевое возбуждение, проецируется на nucl. parafascicularis, centralis lateralis, reticularis (Bowsher, 1961), от которых известны пути, идущие к хвостатому ядру (Walker, 1938, С. Vogt, О. Vogt, 1941; Nauta, Whitlock, 1954).
Наши эксперименты и литературные данные дают основание заключить, что через хвостатые ядра проходит информация, которая играет роль в проведении к коре условного возбуждения, переключающегося в дальнейшем на эффекторные центры оборонительной реакции. Так же, как в отношении ретикулярной формации, нет основания исключить мысль, что активация хвостатых ядер в четвертой стадии образования условного рефлекса связана и с нарастанием возбуждения в эфферентных элементах этого ядра, входящего в экстрапирамидную систему.
Полученные данные не отрицают возможности, что возбуждение, направляющееся в моторные аппараты коры, опосредуется и другими путями (гипоталамическими, субталамическими ядрами и т. д.), которые предстоит еще шире обследовать. Не исключена также возможность, что в локальной активации моторных кортикальных компонент участвуют другие системы связи, в том числе прямые волокна от слухового нерва. Что это вполне вероятный путь, показывают наблюдения Ю. В. Бирючкова (1963) из лаборатории С. Б. Дзугаевой, который нашел прямые связи зрительного нерва с сенсомоторной зоной коры головного мозга. Справедливость подобного предположения подтверждается и наблюдениями В. В. Артемьева и Н. И. Безладновой (1952), которые в сенсомоторной зоне коры мозга кошек в ответ на действие условного звукового сигнала в стадии упроченного рефлекса обнаружили вызванные потенциалы, следующие с латентным периодом, более коротким, чем в первичной проекционной зоне слуха. Эти данные подтверждают предположение, что влияния со слухового анализатора на двигательный осуществляются и с помощью путей с тем же количеством переключений по их ходу, что и тех, которые оканчиваются в корковом конце слухового анализатора. По аналогии со зрительным анализатором (Ю. В. Бирючков, 1963) возможно предположить, что этот путь отходит от слухового анализатора на уровне corona radiata и, следовательно, не опосредуется неспецифическими системами.
В систему путей, участвующих в IV стадии формирования упроченных условных рефлексов, не входит система обонятельного мозга (комплекс миндалевидное ядро - гиппокамп). В этом отношении наши данные подтверждают наблюдения (Yoshii и сотр., 1958а; Н. Иошии и сотр., 1958б; К. Лишак, Э. Граштьян, 1958, и др.), в которых показано, что роль этих структур в формировании временной связи сводится к участию их в начальных этапах выработки условной реакции. Нет основания думать, как это делают Adams и сотр., (1962), что образования обонятельного мозга играют главную роль в функции памяти.
Изменения ЭГ на IV стадии выработки рефлекса начинаются и сопровождаются сдвигами электрической активности на следах условных реакций. Однако эти следовые процессы сохраняются в течение значительно менее длительного времени, чем в слуховом анализаторе на предыдущей стадии выработки рефлекса. Формирование этих следовых реакций указывает на их органическую связь с условнорефлекторным процессом. Следует отметить, что аналогичные картины активности регистрировались и в межсигнальные периоды, а также в ответ на внешние раздражения, что является свидетельством того, что вновь сформировавшаяся констелляция является доминантной.
Таким образом, в IV стадии образования условного оборонительного рефлекса наступает некоторая перестройка прежней доминантной констелляции. По-видимому, уменьшение объема основных и побочных компонент условной реакции приводит к снижению притока возбуждения болевого происхождения к сигнальному (слуховому) анализатору, и условная реакция начинает вызываться при более слабом уровне возбуждения этого анализатора или, возможно, при участии отдельных его звеньев. Новая констелляция выражается в избирательной активации всех компонент кожно-двигательного анализатора и особенно тех структур, которые причастны к проведению и восприятию болевой афферентации (центральное серое вещество, ядра ретикулярной формации ствола мозга, коры сигмовидной извилины, а также хвостатые ядра). Если в III стадии выработки условного оборонительного рефлекса в условнорефлекторную констелляцию входили системы взаимодействующих анализаторов с акцентом на системе сигнального анализатора, то в замыкании на IV стадии участвует та же доминантная констелляция, но акцент ее смещается в сторону безусловнорефлекторной системы, включающей сенсорные и моторные компоненты кожно-двигательного анализатора. Следует отметить, что смещение акцента процессов в сторону безусловнорефлекторного анализатора касается преимущественно его подкорково-стволовых уровней, но это наблюдение требует дальнейшего изучения, прежде чем удастся сделать окончательные выводы о динамике значимого для замыкания взаимодействия процессов. Полученные нами данные позволяют лишь утверждать, что в период формирования условного оборонительного рефлекса происходит изменение отношения афферентного и эфферентного звеньев дуги условного рефлекса. Очевидно, изменениям этих соотношений обязаны разноречивые сведения, которые имеются в литературе по поводу взаимодействия безусловных и условнорефлекторных центров (см. Ф. П. Майоров, 1954). Неслучайно, что выраженное взаимодействие между условной и безусловной реакциями у собак с "задолбленными" рефлексами отсутствует в отличие от собак с "молодыми" условными рефлексами (Р. Л. Винник и Н. Г. Иванова, 1962).
Все исследователи отмечают, что в стадии глубоко упроченных рефлексов изменений электрической активности отметить не удается. Перестают регистрироваться и вызванные ответы в корковом конце сигнального анализатора (В. В. Артемьев, Н. И. Безладнова, 1952). Наши данные показывают, что по мере дальнейшего упрочения рефлекса исчезают отчетливо выраженные сдвиги ЭГ в сенсомоторной зоне коры, в ретикулярных структурах и хвостатых ядрах. Каких-либо локальных процессов в этой стадии выработки рефлекса отметить не удается. Это свидетельствует, видимо, о том, что осуществление условного рефлекса протекает со столь экономичными затратами энергии и на таком уровне, который не представляется возможным обнаружить электроэнцефалографически.
В V стадии формирования условного оборонительного рефлекса на ритмические (2 в 1 сек) звуковые раздражения (щелчки) исчезают и вызванные ответы из коркового конца сигнального (слухового) анализатора (см. рис. 35, б). Сначала исчезают вызванные потенциалы на 2-й, 3-й, 4-й, 5-й и 6-й щелчки, а затем и на 1-й щелчок.
На этой стадии формирования условного рефлекса можно было наблюдать время от времени (в зависимости, очевидно, от функционального состояния животного) появление в ответ на действие условного сигнала картин ЭГ, характерных для одной из предыдущих стадий выработки рефлекса. При длительной работе с собаками удается наблюдать в особенно отчетливой форме (хотя та же закономерность выступает и при менее продолжительных исследованиях), что изменения электрических процессов в сенсомоторной зоне коры выражаются в появлении эффекта десинхронизации вместо наблюдавшейся на первых этапах выработки рефлекса экзальтации электрических колебаний. ЭЭГ сенсомоторной зоны коры в стадии значительно упроченных рефлексов становятся очень сходными с ЭГ всех других зон коры мозга. В этой зоне к концу стадии перестают регистрироваться и вызванные ответы (см. рис. 19). Это свидетельствует об изменении глубины возбудительного процесса, охватывающего и сенсомоторное представительство. Итак, архитектура возбуждения, сопутствующая пятому этапу формирования условного рефлекса, опять меняется - условные звуковые сигналы вызывают очень слабые генерализованные изменения ЭГ, свидетельствующие о диффузном распространении возбуждения, вызываемого сигнальным раздражением.
Иными словами, те локальные процессы, которые определяют осуществление условной реакции, осуществляются столь экономно, что их не представляется возможным обнаружить в условиях электрофизиологической методики (и в этом несомненный недостаток электрофизиологического метода).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'