Глава III. Взаимодействие специфической и неспецифической афферентации и его роль в первый период концентрации (III стадии) условного оборонительного рефлекса
Локальные изменения электрической активности, дающие возможность привести электрографические доказательства участия взаимодействующих анализаторов в замыкании, удалось получить на более поздних стадиях образования рефлекса, когда несколько сглаживались диффузные процессы, маскировавшие эти изменения на первых этапах формирования условной реакции. Фазные сдвиги электрической активности, приходящие на смену генерализованным реакциям, представляют собой отражение тех процессов иррадиации и концентрации в системе анализаторов, вступающих во временную связь, которые были изучены в школе И. П. Павлова по показателям эффекторного условнорефлекторного ответа.
Электрофизиологически явление концентрации процессов было обнаружено в исследованиях М. Н. Ливанова, К. Л. Полякова (1945), М. Н. Ливанова и сотр. (1951), а затем подтверждено и другими исследователями (В. В. Артемьев, Н. И. Безладнова, 1952; В. В. Артемьев, 1959, 1962; Л. Г. Трофимов и сорт., 1955; Р. Н. Лурье и сотр., 1956; О. В. Верзилова, 1955, 1958; Morrell, Naquet, 1956; Т. М. Мохова, 1956; Т. А. Серегина, 1956; Г. Т. Сахиулина, 1957, 1960, 1958; Yoshii, 1957; Gastaut и сотр., 1957; Morrell, 1957; А. Б. Коган, 1960; Chow и сотр., 1957; Morrell и сотр. 1957; Rougeul, 1958; Т. С. Наумова, 1958, 1960, 1961, 1962; Л. Г. Трофимов и сотр., 1958, 1959; С. М. Бабиян, 1960, Jus, Jus, 1959а, б; Т. Б. Швец, 1960, и др.) М. Н. Ливановым и его сотрудниками было выяснено, что период синхронизации активности, совпадающей с периодом генерализации временных связей, постепенно ослабляется. Обычно дело начинается с того, что во взаимодействующих анализаторах наступает большее повышение реактивности, чем в других структурах (В. Н. Думенко, 1955а, б). Во взаимодействующих анализаторах наблюдается и более выраженное нарастание возбуждения (рост ритма и амплитуд колебаний). Так, О. В. Верзилова (1955), Р. Н. Лурье и сотр. (1956), В. Н. Думенко (1955а, б, 1960а, б) и другие показали, что при упрочении оборонительных условных рефлексов у собак генерализованные по коре изменения электрической активности сменяются концентрацией процессов в корковых концах анализаторов, объединяющихся во временную связь. Это выражается в нарастании электрической активности (по частотно-амплитудным показателям) и синхронизацией ритмики потенциалов в этих зонах. В топоскопических исследованиях это выражалось в появлении фокусов активности (МВКП), преимущественно приуроченных к кортикальным уровням взаимодействующих анализаторов (М. Н. Ливанов, 1958). Эти наблюдения привели к предположению, что МВКП формируются главным образом в тех нервных структурах, которые в связи с выработкой условного рефлекса приходят в доминантное состояние. При формировании оборонительного условного рефлекса у собак наибольшее нарастание возбуждения наступает в слоях II, IV коркового конца анализатора, воспринимающего условный сигнал, и в слое V двигательного анализатора (М. Я. Рабинович, 1958, 1961). Если сигнал оборонительной реакции носит ритмический характер, эти ритмы у собак также приурочиваются к взаимодействующим анализаторам (слуховому и двигательному) (О. В. Верзилова, 1955). Наблюдение преимущественного вовлечения в реакцию корковых концов анализаторов, участвующих в формировании рефлексов, выступило и в исследованиях на людях по оборонительной (Т. А. Серегина, 1956) и речедвигательной (Т. М. Мохова, 1956) методикам. Концентрация процессов в анализаторе условного сигнала обнаружена и по показателю медленной электрической активности (Т. Б. Швец, 1960).
Описываемая стадия выработки рефлекса может быть обозначена как первая стадия концентрации в противоположность той стадии концентрации (можно назвать ее второй), которую описали М. Н. Ливанов и К. Л. Поляков (1945) и которая выражается в концентрации ритмических колебаний потенциала в сенсомоторной зоне коры мозга кролика.
Какие же этим изменениям сопутствуют сдвиги электрической активности на уровне субкортикально-стволовых структур?
Как отмечалось, в первой и во второй стадиях выработки условного рефлекса прекращение совместного действия раздражителей сопровождается остаточной активацией. Последняя имеет диффузный характер, однако по мере сочетаний все больше приурочивается к системам взаимодействующих анализаторов, что является предвестником III фазы выработки рефлекса - периода концентрации процессов в слуховом и двигательном анализаторах. Наиболее выраженные изменения электрической активности на следах сочетаний отмечаются в системе слухового анализатора. На рис. 29 представлены сдвиги ЭГ корковых и бульбарных структур собаки № 11, сопровождающие условную реакцию на II (см. рис. 29, а) и в начале III этапа (см. рис. 29, б) образования рефлекса. В ответ на 42-е применение подпорогового тона 500 гц 40 дб произошло возрастание фоновой активности, которое было в одинаковой степени выражено в кохлеарных ядрах, гигантоклеточном ядре ретикулярной формации, средней ножке мозжечка, а также в менее отчетливой форме, но все-таки выступало в корковых зонах мозга. Экзальтация активности бульбарных структур наступила с латентным периодом 150 мсек, и почти одновременно с ней тот же процесс возник в корковых зонах мозга. Болевое подкрепление вызвало усиление электрических колебаний в кохлеарных ядрах и коре сильвиевой зоны, которое сохранялось и в последействии.
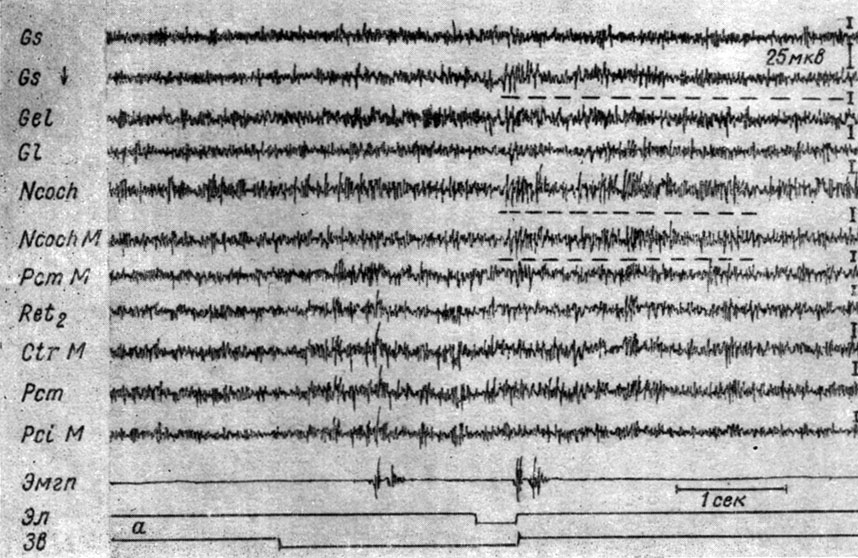
Рис. 29. Следовые изменения ЭГ (подчеркнуты пунктирной линией) в системе слухового анализатора (кохлеарных ядрах, сильвиевой извилине коры большого мозга) собаки № 11, наступающие во II (а) и в начале III (б) стадии формирования условного оборонительного рефлекса. а - 42-е и б - 53-е сочетания тона 500 гц 40 дб с электрокожным раздражением правой передней лапы (а - опыт № 36 от 4/VIII 1960 г.; б - опыт № 49 от 23/VIII 1960 г.)
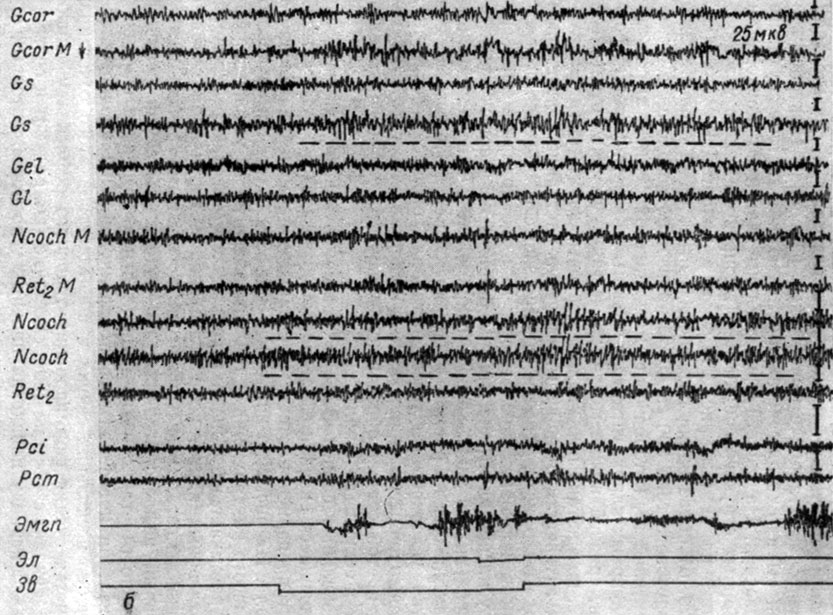
Рис. 29. Продолжение
Как следует из рис. 30, продолжительность этого этапа выработки условного рефлекса составляла около 46 сочетаний. Следовые процессы, приуроченные к системе слухового анализатора, в этой стадии образования условного рефлекса длились десятки секунд (до 1 мин).
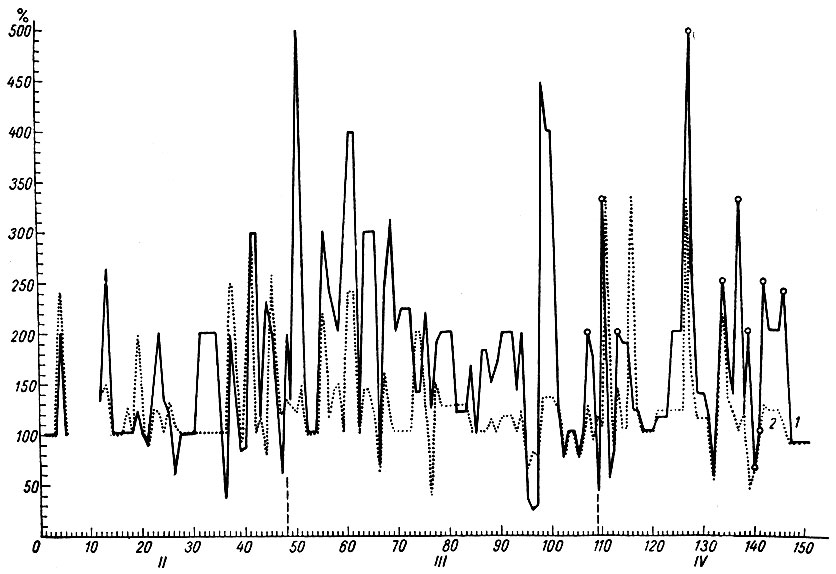
Рис. 30. Амплитудная характеристика ЭГ условной реакции при формировании оборонительного рефлекса на тон 500 гц 40 дб (собака № 11). 1 - ЭГ кохлеарного ядра; 2 - ЭГ ретикулярной формации продолговатого мозга (гигантоклеточное ядро). Кружки - амплитуда медленных колебаний. По оси абсцисс - порядковые номера сочетаний и стадии формирования рефлекса; по оси ординат - проценты по отношению к исходным величинам
Локальные сдвиги ЭЭГ в системе слухового анализатора, появлявшиеся сначала в последействии рефлексов, в III стадии регистрировались в ответ на очередное применение условного сигнала. На рис. 29, б видно, что при 53-м сочетании звукового и болевого раздражения у той же собаки № 11 условный рефлекс протекает на фоне активации электрических осцилляций в кохлеарных ядрах, коре сильвиевой и коронарной извилин большого мозга (регистрируемых погружными электродами). Условный рефлекс возникает на фоне описываемой реакции. Особенно отчетливо видна избирательная активация слухового анализатора после болевого подкрепления. Она продолжалась в течение нескольких секунд в последействии этого сочетания.
Третья стадия формирования условного рефлекса у собаки № 11 длилась с 47-го по 109-е сочетание (см. рис. 30) и соответствовала периоду появления прочных условнорефлекторных ответов, латентный период которых к концу стадии значительно снизился. Изменения активности в системе слухового анализатора были особенно четко выражены в начале третьего этапа. К концу этого периода (см. рис. 30) они значительно уменьшились.
Приведенные наблюдения свидетельствуют о том, что в III стадии образования рефлекса в системе слухового анализатора происходит избирательное нарастание уровня возбуждения, которое способствует осуществлению условного рефлекса и его последействию. Параллельно с этим возрастание возбуждения происходит и в корковом конце кожно-двигательного анализатора. Особенно четко это выступило в экспериментах с собакой № 4. На рис. 31 представлено 123-е применение порогового звукового раздражения (тон 500 гц 48 дб), ранее сочетавшееся с пороговым электрокожным раздражением у этой собаки. Видно, что вместо генерализованного усиления электрических колебаний (см. рис. 25) наступила их локальная активация в кохлеарных ядрах, в сенсомоторной зоне коры (более чем вдвое по сравнению с фоном). В остальных корковых территориях наблюдается лишь возрастание частоты изменений потенциала без существенных сдвигов в амплитудном показателе. Усиление возбуждения во взаимодействующих анализаторах появляется с латентным периодом 1/2 сек и продолжается весь период действия звука, затягиваясь в последействии на 3-5 сек, что особенно отчетливо выступает в кохлеарных ядрах. Заметного нарастания возбуждения в корковом конце слухового анализатора отметить не удается, так как запись ЭЭГ эктосильвиевой извилины произведена накладными электродами.
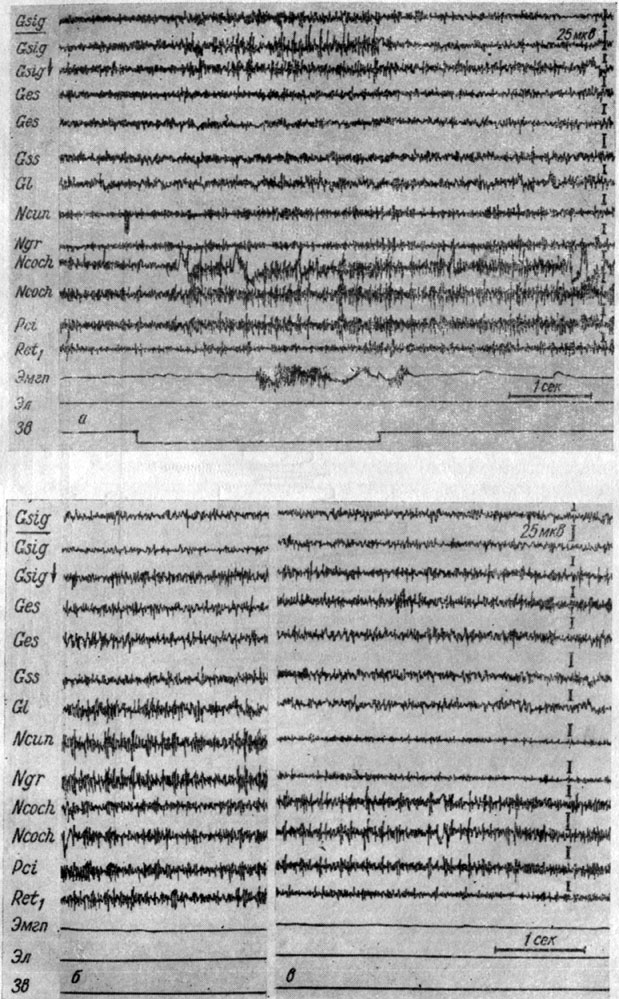
Рис. 31. Изменения ЭГ структур коры большого мозга и продолговатого мозга собаки № 4, наступающие в III стадии формирования условного оборонительного рефлекса (а), а также в межсигнальные периоды (б, в), а - 123-е применение тона 500 гц 48 дб - сигнала оборонительной реакции (опыт № 56 от 8/IV 1959 г.; б - опыт № 54 от 3/IV 1959 г.; в - опыт № 56 от 8/IV 1959 г.)
Третья фаза образования условного рефлекса на тон 500 гц 48 дб у собаки № 4 длилась с 108-го по 170-е сочетание (см. рис. 15, а, б). В этот период окончательно установились регулярные условнорефлекторные реакции (в 100% случаев) со стабильным, снижением по сравнению с первыми стадиями, латентным периодом.
Сопоставление характера условнорефлекторных изменений ЭЭГ бульбарных структур (см. рис. 15, а) и величины вызванных ответов (см. рис. 16) в III стадии формирования условного оборонительного рефлекса на включение условного сигнала (тона 500 гц 48 дб) у собаки № 4 показало, что в начале этого периода происходит значительное увеличение амплитуды вызванных потенциалов, которая к концу третьей стадии снижается. Обращает на себя внимание, что к концу III стадии обнаруживается тенденция к снижению степени активации электрических процессов в системе слухового анализатора (кохлеарных ядрах) и нарастанию ее в системе кожно-двигательного анализатора (сенсомоторной зоне коры). Это проявляется как в отношении изменений электрической активности, сопровождающих условные рефлексы (см. рис. 15, а, б), так и в отношении следовых процессов (см. рис. 17, а, б).
Сохранение следов условнорефлекторного возбуждения в III фазе формирования условного рефлекса, наблюдавшееся в системе взаимодействующих анализаторов и в первую очередь в кохлеарных ядрах и сенсомоторной зоне коры, длилось от десятков секунд до 2 мин (см. рис. 18).
На следах рефлексов в III стадии их выработки наблюдается явление избирательного активирования преимущественно кохлеарных ядер, однако его можно видеть и в других системах мозга. Вот некоторые иллюстрации сказанного. На рис. 31, б, в представлены ЭГ коры и ядер продолговатого мозга собаки № 4, записанные в разные межсигнальные периоды. В первом (см. рис. 31, б) наблюдается явление преимущественного возрастания активности ядер нежного и клиновидного канатиков, во втором (см. рис. 31, в) - активация в кохлеарных ядрах и выраженная депрессия ее в ядрах нежного и клиновидного канатиков и нижнем центральном ядре ретикулярной формации. В корковых структурах существенных различий в этих записях ЭГ отметить не удается в связи с тем, что ЭГ этих формаций регистрировалась с помощью накладных электродов.
В тех случаях, когда отведение электрических потенциалов происходило погружными электродами, следовые изменения электрической активности кортикальных компонент слухового анализатора были четко выражены.
Как следует из приведенных выше наблюдений, при осуществлении условных оборонительных рефлексов на III стадии их формирования одновременно с локальной экзальтацией электрических процессов в системе слухового и кортикальных компонент кожно-двигательного анализаторов в других структурах мозга происходит слабый диффузный эффект активации. Однако у некоторых животных в подкорково-стволовых образованиях, не принадлежащих к сигнальному анализатору, наступало падение активности, которое охватывало и стволовые звенья кожно-двигательного анализатора (в частности, ядра нежного и клиновидного канатиков). Сказанное иллюстрирует рис. 26, б. Видно, что у собаки № 3 83-е применение условного звукового сигнала сначала вызывает снижение активности в структурах продолговатого мозга и эффект десинхронизации в корковых концах слухового и двигательного анализаторов. К моменту оборонительной реакции появляется еще большее снижение активности в дугообразных волокнах, идущих от ядер нежного и клиновидного канатиков (до 5-10 мкв). В ряде отведений от кохлеарного ядра амплитуды колебаний, напротив, увеличиваются (до 30-40 мкв). Весь период действия звукового сигнала и в последействии от него в зрительной зоне наблюдаются высокоамплитудные отклонения, аналогичные тем, которые возникают при действии светового раздражения. В стадии концентрации высокоамплитудные отклонения в зрительной зоне коры также отчетливо выражены, как и во II стадии генерализации (см. рис. 26, а). В то же время объем диффузных процессов на уровне ствола по мере упрочения рефлекса несколько снизился.
Измерения величины вызванных ответов, зарегистрированных в ЭЭГ кортикальных формаций на включение условного сигнала (тона 500 гц 48 дб) у собаки № 3 (см. рис. 19), показали, что в третьей стадии формирования условного рефлекса происходит значительное увеличение амплитуды вызванных потенциалов, регистрируемых в коре эктосильвиевой извилины. К концу третьего периода выработки рефлексов величина этих ответов снижается. В период наибольшего возрастания вызванных потенциалов в коре эктосильвиевой извилины резко уменьшаются ответы в коре сигмовидной извилины, а в конце III периода формирования рефлекса, когда снижаются ответы в первичной слуховой проекционной зоне, наоборот, резко возрастают вызванные потенциалы в сенсомоторной зоне коры. Запись активности кортикальных формаций производилась накладными электродами.
Преобладание на третьей стадии образования рефлекса локальных изменений ЭГ слухового анализатора наблюдается и в его среднемозговых звеньях. Так, на рис. 32 видно, что вместо генерализованной экзальтации (см. рис. 27) при 141-м применении условного сигнала (тона 500 гц 48 дб) у собаки № 2 происходит возрастание электрической активности внутреннего коленчатого тела и в меньшей степени коры супрасильвиевой извилины. В заднем двухолмии, ретикулярной формации среднего мозга, центральном сером веществе та же тенденция выражена значительно слабее. В коре коронарной извилины нарастание частоты колебаний сопровождается уменьшением их величины. Моторная реакция (слабые токи действия) появляется на третьей секунде. Электрокожное раздражение сопровождалось эффектом генерализованной депрессии активности.
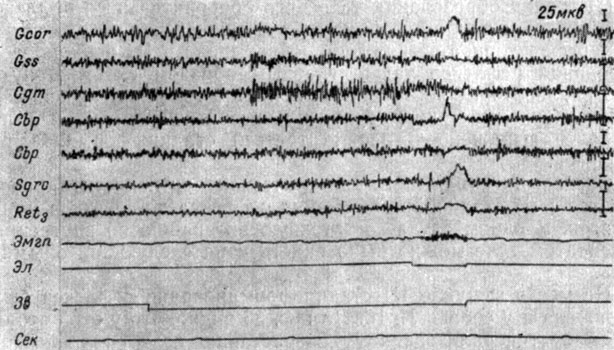
Рис. 32. Изменения ЭГ коры большого мозга и структур среднего мозга собаки № 2, наступающие в III стадии формирования оборонительного условного рефлекса. 141-е сочетание звукового сигнала (тон 500 гц 48 дб) с электрокожным раздражением (опыт № 57 от 27/XII 1958 г.)
III стадия образования условного оборонительного рефлекса у собаки № 2 длилась с 47-го по 148-е сочетание. Следовое условнорефлекторное возбуждение охватывало в III периоде формирования рефлекса также главным образом систему слухового анализатора (внутреннее коленчатое тело, в меньшей мере заднее двухолмие, кору супрасильвиевой извилины), и было наиболее выражено в начале третьей стадии формирования рефлекса.
Следовые изменения электрической активности продолжаются в III стадии формирования рефлекса до десятков минут и часов. В одном случае остаточную активацию во внутреннем коленчатом теле можно было видеть и на следующий опытный день.
Не менее выпукло III стадия формирования рефлекса была выражена и у собаки № 16. На рис. 33 видно, что у этой собаки на смену генерализованному возрастанию колебаний электрических потенциалов коры, внутреннего коленчатого тела, заднего двухолмия, красного ядра, центрального серого вещества (см. рис. 33, а), которое регистрировалось в течение 60 первых сочетаний (I электрографическая фаза выработки рефлекса у этого животного отсутствовала), приходят более локальные процессы (см. рис. 33, б). На рис. 33, б представлено 62-е воздействие звукового сигнала, подкрепляемого электрокожным раздражением. Тон сопровождается значительным ростом частоты изменений потенциалов внутреннего коленчатого тела с 45-50 до 57-62 в 1 сек и их амплитуд с 15-20 до 40-60 мкв. Это совпадает с несравненно меньшим увеличением тех же показателей в остальных мезенцефалических структурах, как-то: заднем двухолмии, ретикулярной формации, красном ядре, а также зрительной, слуховой и сенсомоторной зонах коры. Эта электрографическая картина сохраняется и в последействии (1 сек). По мере сочетаний ЭГ внутреннего коленчатого тела все более отличалась от ЭЭГ смежных областей мозга, и описываемые изменения усиливались и сохранялись по нескольку секунд в последействии сочетания. Четкой корреляции эффекта активации коленчатого тела и сдвигов ЭЭГ слуховой зоны коры здесь не удалось отметить вследствие записи ее потенциалов накладными электродами. Продолжительность III стадии формирования условного рефлекса у этой собаки - с 61-го по 165-е сочетание.
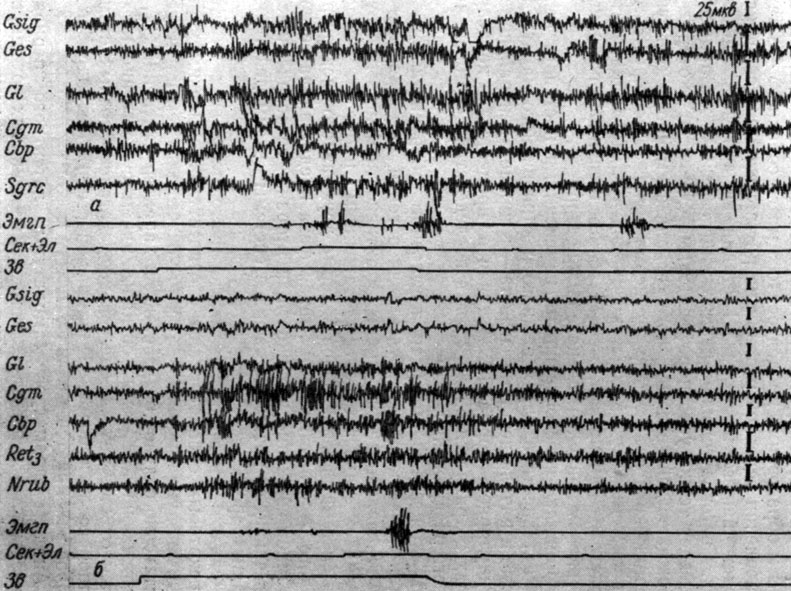
Рис. 33. Изменения ЭГ коры большого мозга и образований среднего мозга собаки № 16, наступающие во II (а) и III (б) стадиях формирования условного оборонительного рефлекса. а - 46-е, б - 62-е применение условного сигнала (тон 500 гц 48 дб). а - опыт № 20 от 20/I 1958 г.; б - опыт № 24 от 4/11 1958 г. (по Т. С. Наумовой, 1961)
Следовое возбуждение может проявляться не только сразу после прекращения сочетания стимулов, но также в периоды, значительно отставленные от условных рефлексов.
Констелляция процессов, формирующаяся в III стадии образования рефлекса, имеет доминантные свойства, она воспроизводится болевыми, световыми и другими раздражениями, а также выступает при выработке условной реакции на другие, например на световые, стимулы. На рис. 34 представлены записи ЭГ коры и образований продолговатого мозга собаки № 4 в начале становления условной реакции на ритмический световой сигнал, происходившего на фоне III стадии выработки рефлекса на звук. Световое условное раздражение с первых же применений вызывает те же условнорефлекторные сдвиги ЭГ, которые сопровождают действие звукового сигнала, а именно локальную активацию кохлеарных ядер и сенсомоторной зоны коры. Активация системы слухового и кожно-двигательного анализаторов происходит одновременно с появлением вызванных потенциалов в корковом конце зрительного анализатора.
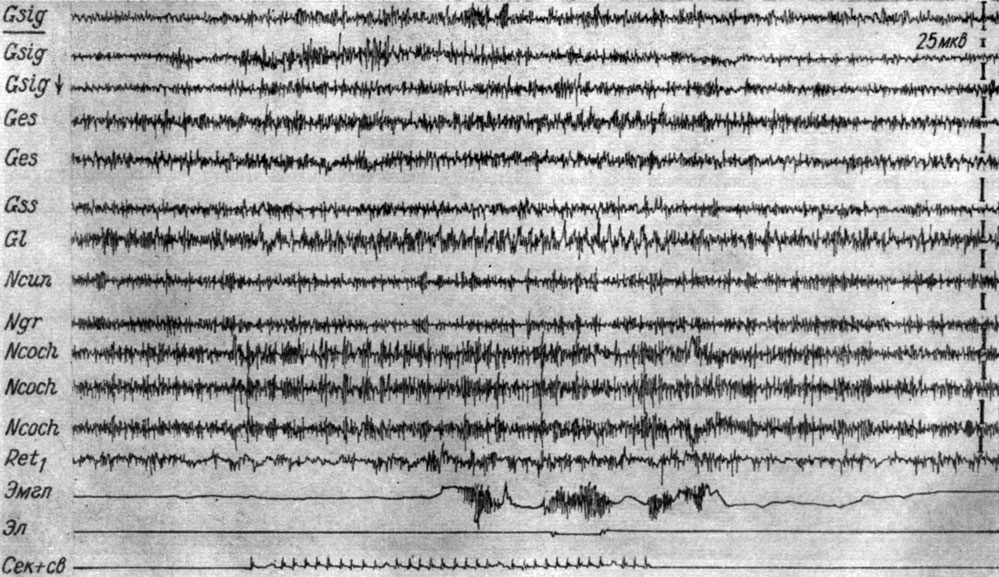
Рис. 34. Изменения ЭГ коры большого мозга и образований продолговатого мозга собаки № 4 в начале формирования условного оборонительного рефлекса (6-е сочетание) на вспышку света (0,25 лк, 10 в 1 сек). Формирование рефлекса начато на фоне III стадии выработки условного оборонительного рефлекса на звук (опыт № 59 от 11/IV 1959 г.)
Параллельно с описанными изменениями наступает депрессия активности в ядрах нежного и клиновидного канатиков, а также в начале действия сигнала - в нижнем центральном ядре ретикулярной формации. Активация процессов в кохлеарных ядрах сохраняется около 2 сек в последействии светового и болевого раздражений.
В процессе формирования условного оборонительного рефлекса на звуковые стимулы надпороговой интенсивности стадия генерализованного снижения активности мозговых структур сменялась стадией с более выраженными процессами в основных системах анализаторов в форме двухфазного электрографического ответа, в котором первая фаза представляла генерализованное снижение активности, а вторая - ее возрастание, охватывающее преимущественно системы взаимодействующих анализаторов.
Однако такое явление имело место не у всех животных. У некоторых собак сигналы с относительно высоким уровнем энергии звука (например, 100 дб) оказывали, тем не менее, недостаточно сильное воздействие, и характер изменений ЭГ, сопровождающих условные сигналы, ничем существенно не отличался от динамики процессов на действие условных стимулов подпороговой и пороговой интенсивности. Более подробно этот вопрос разбирается в главе VI.
Анализ параметров (амплитуды, латентного периода и длительности) первых трех компонент вызванных потенциалов в первичной проекционной зоне слуха, регистрируемых с помощью накладных макроэлектродов в ответ на условные ритмические (2 в 1 сек) звуковые раздражения (щелчки) у собак показал, что в III стадии формирования условного оборонительного рефлекса происходит усиление тех сдвигов, которые наблюдались в предыдущий период выработки рефлекса, а именно величина всех компонент ответа на первый щелчок (или два первых щелчка) еще более возрастает. Сказанное иллюстрируют рис. 35, а также рис. 22. Как следует из этих рисунков, в III фазе образования рефлекса происходит дальнейшее (см. рис. 22) увеличение и первой позитивной, и негативной, и второй позитивной компоненты ответа на первый щелчок (на графиках - непрерывная линия). Одновременно с увеличением амплитуды каждой компоненты вызванного потенциала на первый щелчок происходит увеличение латентного периода начала и времени до максимума позитивной фазы, а также максимума негативной фазы, которые к концу стадии достигают исходной величины ответа (см. рис. 23). Увеличение латентного периода позитивной и негативной компоненты ответа на первые щелчки происходит параллельно с увеличением их длительности (см. рис. 24).
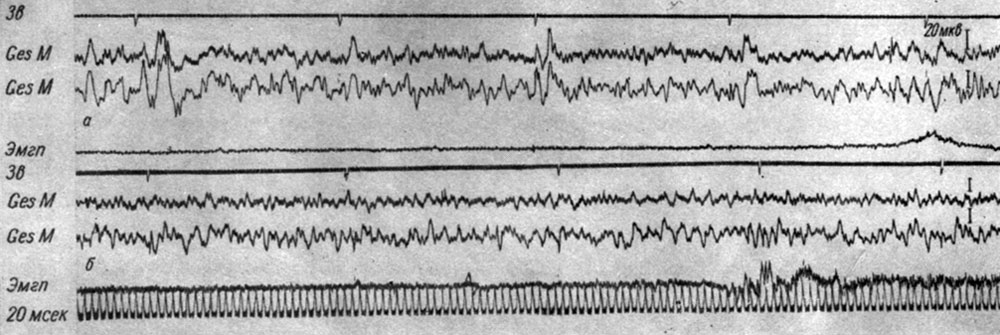
Рис. 35. Изменения вызванных потенциалов в разных участках первичной проекционной корковой слуховой зоны собаки № 19 на III (а) и IV (б) стадиях образований оборонительного условного рефлекса на ритмические звуковые раздражения (щелчки 2 в 1 сек, 56 дб). а - 27-е сочетание (опыт № 12 от 20/V 1964 г.); б - III-е сочетание (опыт № 43 от 18/VI 1964 г.) (по Т. С. Наумовой, Н. С. Поповой и Н. Н. Ульяниной 1966)
По мере приближения к моменту условной реакции амплитуды всех трех компонент первичного ответа значительно уменьшаются (рис. 35, а), нередко до полного их исчезновения (см. рис. 22, 2 и 3). Уменьшение величины каждого из трех компонент ответа к моменту движения конечности происходит параллельно с укорочением их латентного периода. Наиболее ярко это явление выступало для первых двух компонент вызванного ответа у собаки № 20. На рис. 36 представлено графическое выражение изменений латентного периода (отложены по оси ординат) вызванных потенциалов на каждый из 6 щелчков (отложены по оси абсцисс) в 8 опытах, взятых из разных периодов образования условного рефлекса у собаки № 20. Видно, что до 146-го сочетания двигательной условной реакции (отмечено стрелкой) предшествует или сопутствует период снижения латентного периода вызванных ответов.
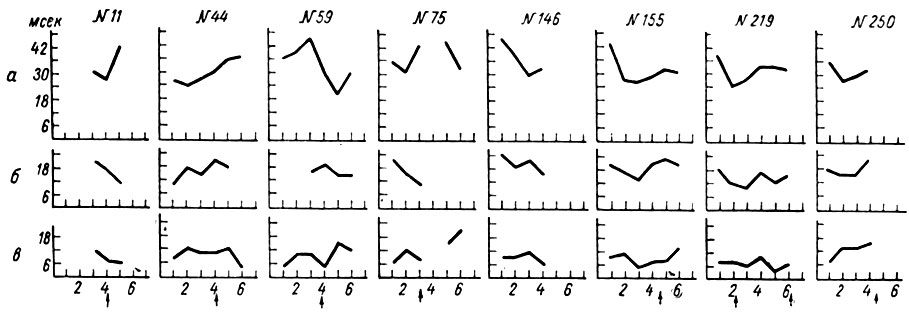
Рис. 36. Динамика латентного периода вызванных ответов на разных стадиях формирования оборонительного условного рефлекса у собаки № 20. а, б, в - то же, что на рис. 23. По абсцисс - порядковые номера щелчков; по оси ординат - миллисекунды. Сверху обозначены номера сочетаний. Стрелки - условнорефлекторные реакции (по Т. С. Наумовой и Н. С. Поповой, 1967а)
Длительность фаз вызванного ответа, так же как их латентный период и амплитуда, существенно меняются в момент, предшествующий и сопутствующий условнорефлекторной реакции конечности, а именно в первых трех стадиях формирования рефлекса условнорефлекторная реакция сопровождалась сокращением длительности ответов. Так, на рис. 37 представлены ЭЭГ коры эктосильвиевой извилины большого мозга собаки № 19, записанные в разные периоды формирования у нее условного оборонительного рефлекса. Видно, что в каждый период образования условной реакции (а, б, в) существенно различаются вызванные потенциалы, появляющиеся в ответ на первый и каждый из последующих щелчков. На рис. 37, а видно, что длительность вызванных потенциалов существенно меняется от первого к последующим щелчкам даже тогда, когда еще отсутствует условнорефлекторная реакция конечности (6-е сочетание); в III стадии образования рефлекса (рис. 37, б) длительность всех компонент вызванного потенциала на 2-й щелчок, совпадающий с условнорефлекторным двигательным ответом, значительно укорачивается по сравнению с ответом на первый щелчок. Ответы на 3-й, 4-й и 5-й щелчки по длительности приближаются к реакции на 1-й щелчок. В первых трех стадиях укорочение длительности вызванных ответов в момент, предшествующий и сопровождающий движение конечности (т. е. на 3-й и 4-й щелчки), отчетливо видно и на рис. 24.
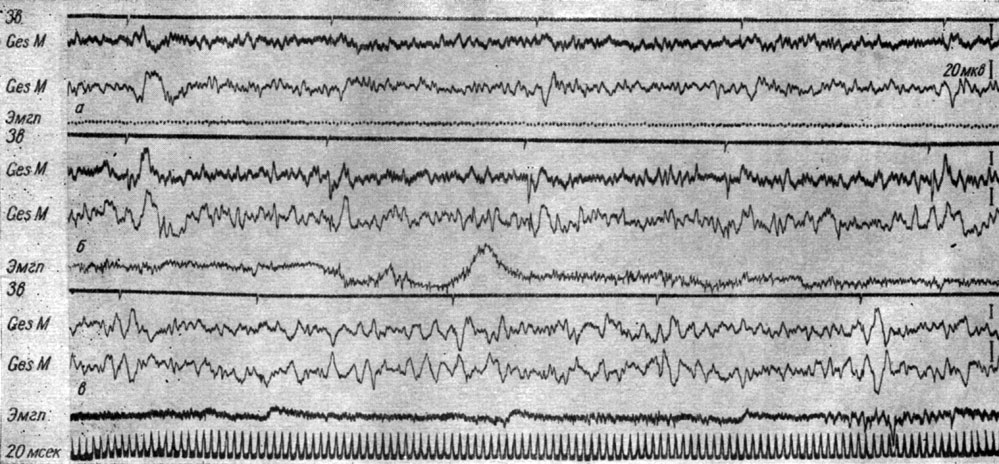
Рис. 37. Изменения вызванных потенциалов в разных участках первичной проекционной корковой слуховой зоны собаки № 19 на I (а), III (б) и IV (в) стадиях образования оборонительного условного рефлекса на ритмические звуковые раздражения (щелчки 56 дб, 2 в 1 сек). а - 6-е сочетание (опыт № 3 от 6/V 1963 г.); б - 29-е сочетание (опыт № 24 от 18/VI 1963 г); в - 162-е сочетание (опыт № 58 от 29/IX 1963 г.) (по Т. С. Наумовой и Н. С. Поповой, 1967а)
В III стадии формирования условного оборонительного рефлекса происходит стабилизация латентного периода условнорефлекторного ответа, который все более приближается к моменту подкрепления (см. рис. 22, а).
Введение дифференцировки приводит к изменениям не только амплитуды, но также латентного периода и длительности всех компонент вызванного ответа, а именно введение дифференцировки всегда (см. рис. 22) сопровождается увеличением амплитуды ответа, а также укорочением латентного периода реакции (см. рис. 23) на щелчки, служащие сигналом условной оборонительной реакции. Аналогичным образом сказывается введение дифференцировки и на длительности вызванных ответов, которая также уменьшается (см. рис. 24). После упрочения дифференцировки восстанавливается динамика изменений амплитуды, латентного периода и длительности первых фаз ответа, имевшая место до введения дифференцировки.
Итак, условная реакция на звуковой сигнал подпороговой и пороговой интенсивности на III этапе выработки рефлекса формируется на фоне локальной активации (возрастания частоты и амплитуд колебаний) электрической активности, обнаруженной в записях погружными электродами на разных уровнях слуховой системы (кохлеарных ядрах, трапециевидном теле, верхней оливе, внутреннем коленчатом теле, коре сильвиевой, эктосильвиевой и супрасильвиевой извилин) и корковом конце кожно-двигательного анализатора (сигмовидная извилина).
Возникновению активации в системе взаимодействующих анализаторов предшествует избирательное возрастание электрической активности слуховой системы, появляющееся сначала на следах условных рефлексов во II стадии их формирования, а затем и в III стадии.
В III период выработки рефлекса обнаруживается четкая корреляция электрических реакций на разных уровнях слухового анализатора как в период действия сигнальной стимуляции, так и непосредственно на следах сочетаний и в межсигнальные периоды. Видимое несходство изменений корковых и субкортикальных звеньев слухового анализатора у отдельных животных было связано с неадекватным способом отведения потенциалов от этих структур. Регистрация ЭЭГ слуховой зоны коры у таких животных проводилась накладными электродами, и в этих записях проявлялся эффект десинхронизации - снижение амплитуд и возрастание ритма электрических колебаний. Включение условного сигнала сопровождается появлением в слуховой зоне коры вызванных ответов, по величине превышающих те, которые имели место во II стадии формирования рефлекса.
При образовании условных оборонительных рефлексов на ритмические (2 в 1 сек) звуковые раздражения (щелчки) в III стадии их формирования происходит дальнейшее увеличение амплитуды, некоторое увеличение латентного периода (который все-таки остается ниже исходных значений) и длительности позитивной, негативной и второй позитивной компоненты вызванных ответов, регистрируемых с помощью накладных электродов в первичной проекционной слуховой зоне на первые щелчки из 6, представляющих сигнал оборонительной реакции.
В противоположность увеличению первичных вызванных потенциалов на первые щелчки в серии по мере приближения к моменту условной реакции происходит их значительная редукция. Это уменьшение идет параллельно укорочению латентного периода и длительности первичных ответов.
Параллельно со значительным нарастанием уровня возбуждения в слуховом и двигательном анализаторах как в момент осуществления условного рефлекса, так и на его следах (последнее касается главным образом слуховой системы, в которой остаточная активация может длиться многие часы) наблюдается значительно менее выраженное усиление электрических процессов (диффузный эффект активации) или иногда их депрессия в ретикулярной формации ствола мозга, ядрах нежного и клиновидного канатиков, в ядрах тройничного нерва, средних ножках мозжечка, вентральном и ретикулярном ядрах таламуса, субталамусе, гипоталамусе, хвостатом ядре, гиппокампе, миндалевидном ядре и т. д. Нивелируются изменения ЭЭГ и в незаинтересованных территориях коры. Таким образом, изменения электрических осцилляций, проявляющиеся в форме некоторого нарастания уровня возбуждения в структурах, непосредственно не заинтересованных в осуществлении условного рефлекса, сопровождают процесс замыкания и на третьем этапе выработки временной связи. Это явление наблюдается как на уровне ствола мозга, в частности ретикулярной формации, так и на уровне корковых структур. В последних это проявляется не только в эффекте десинхронизации, но также в появлении у ряда животных в ответ на условные звуковые стимулы локальных двухфазных высокоамплитудных отклонений в зрительной зоне коры, сходных с первичными вызванными потенциалами.
Локальные и диффузные сдвиги электрической активности корковых и стволово-подкорковых структур в значительной мере зависят от физических свойств сочетаемых раздражений. При выработке условных оборонительных рефлексов на звуковые сигналы надпороговой интенсивности стадия концентрации электрических процессов в системе взаимодействующих анализаторов никогда не бывает в такой мере полной, как на действие сигналов пороговой интенсивности. При действии условных звуковых раздражений, по интенсивности значительно превышающих пороговые значения, наступает сначала генерализованное снижение уровня электрических процессов, за которым следует локальная активация в системе анализаторов, вступающих во временную связь. Таким образом, осуществление оборонительных условных рефлексов на звуковые раздражения, по интенсивности значительно превышающие пороговые значения, происходит при более широком охвате структур мозга возбуждением и, следовательно, в доминантную констелляцию в этом случае входит более широкий круг образований мозга.
Электрографическая картина активности, сопровождающая действие условного звукового сигнала и его последействия, воспроизводится в ответ на посторонние раздражения. Если в III стадии формирования условного рефлекса на звук начать образование оборонительной временной связи на световые сигналы, последние, кроме специфических процессов (вызванных потенциалов в зрительном анализаторе), провоцируют те изменения, которые сопровождают действие звукового раздражения - избирательную активацию слухового анализатора (начиная с кохлеарных ядер и кончая корой эктосильвиевой и сильвиевой извилин), а также сенсомоторной зоны коры. Это явление свидетельствует о доминантных свойствах той констелляции, которая создается на III стадии формирования рефлекса - стадии концентрации процессов в системе взаимодействующих анализаторов.
Локальные изменения электрической активности на уровне стволово-подкорковых структур удается видеть лишь при использовании биполярного способа отведения. При регистрации электрических потенциалов униполярным методом обнаруживается генерализованная экзальтация электрических колебаний.
К концу III стадии появляется тенденция к снижению степени активации электрических процессов в системе слухового анализатора и нарастанию ее в системе кожно-двигательного анализатора, обнаруживающаяся как по показателю суммарной электрической активности, так и по показателю вызванных ответов.
Как уже отмечалось, одновременное появление однозначных сдвигов электрической активности в различных структурах каждого уровня мозга при осуществлении условных рефлексов свидетельствует о том, что процесс замыкания начинается с множественных одновременных взаимодействий процессов возбуждения, вызванных условным и безусловным раздражениями. В этом смысле нельзя не согласиться с теми исследователями, которые предполагают, что взаимодействие афферентаций при замыкании происходит на разных уровнях мозга (А. Фессар, 1958; Fessard, Gastaut, 1958; Н. Ю. Беленков, 1959, 1960, 1966; Л. Г. Воронин, 1966, и др.), ибо каждый условный рефлекс есть результат синтеза безусловных реакций, каждая из которых представляет многоветвистую структуру (Э. А. Асратян, 1953).
Взаимодействие процессов, по-видимому, осуществляется ранее в нижних отделах мозга, чем в верхних. В предыдущей главе уже шла речь о том, что рассматривать полученные наблюдения как свидетельство равнозначности процессов взаимодействия в разных формациях мозга, по-видимому, нельзя, так как, по мере применения условного сигнала, постепенно начинают выступать те локальные процессы, которые были скрыты диффузными изменениями ЭЭГ на первых этапах формирования временной связи. Именно очаговые процессы связывают (М. Н. Ливанов, 1960; Е. Н. Соколов, 1960)с реализацией условного рефлекса. Так как на смену диффузным приходят локальные сдвиги электрической активности, говорящие о том, что по мере выработки рефлекса происходит постепенное сужение полей взаимодействия возбуждений, синтезирующихся во временной связи, процесс замыкания нельзя рассматривать как процесс параллельного сосуществования независимых взаимодействий. Последние составляют звенья цепи, которые находятся в тесной взаимосвязи и взаимозависимости.
Взаимовлияние возбуждений безусловного и условнорефлекторного происхождения оказывается наиболее сильным не в ретикулярной формации, как полагали Yoshii и сотр. (1956), Hernandez-Peon (1957), Gastaut (1955), Г. Гасто (1958), А. Гасто, А. Роже (1958), А. Фессар (1958) и др., и не в межуточных (теменных) структурах коры мозга, как считают Г. Джаспер и сотр. (1958), А. С. Батуев (1966), а в самих взаимодействующих (условном и безусловном) анализаторах, ибо в них имеются наибольшие условия для этого - наибольшая активация специфической для каждого из них афферентацией и дополнительной неспецифической импульсацией (для слухового анализатора - болевой, для двигательного - слуховой). Таким образом, обе специфические системы находятся в отношении взаимодействия процессов в особых условиях. Какие факты говорят в пользу такого предположения? В первую очередь наблюдения павловской школы. Что касается электрофизиологических исследований, то показано, что в процессе выработки рефлекса несколько уменьшаются диффузные процессы и выявляются преимущественные сдвиги электрической активности (ее усиление) в системе взаимодействующих анализаторов (слуховом и двигательном). В отношении процессов в кортикальных компонентах рефлексов это было замечено М. Н. Ливановым и К. Л. Поляковым (1945), М. Н. Ливановым и сотр. (1951), М. Н. Ливановым (1958, 1960), Р. Н. Лурье и сотр. (1956), О. В. Верзиловой (1958), Т. М. Моховой (1956), Morrell (1957), Г. И. Шульгиной (1961) и др.
Аналогичное явление нам удалось обнаружить и при обследовании электрических процессов стволово-подкоркового уровня. Выяснилось, что по мере формирования условной оборонительной реакции на звук, и именно в третьей его стадии, на смену генерализованным сдвигам электрической активности, одновременно наступающим в структурах каждого уровня мозга, приходят более локальные процессы - процессы преимущественной активации субкортикальных, стволовых отделов сигнального (слухового) анализатора (кохлеарных ядер, трапециевидного тела, верхнего оливарного комплекса ядер, заднего двухолмия и внутреннего коленчатого тела. Естественно, этот процесс происходит параллельно с активацией корковых компонент условного рефлекса.
Одновременно с нами (Т. С. Наумова, 1958) этот факт был показан в отношении условных рефлексов на ритмические световые раздражения у кошек (Г. В. Мегун, 1958). Наблюдения постепенной концентрации вызванных ответов в системе сигнального анализатора подтвердили John и Кillam (1959) на примере оборонительного рефлекса на прерывистый свет у кошек и Chow (1960) при выработке пищевых условных реакций на световые раздражения у обезьян. По данным McAdam и сотр. (1961), в первую очередь сглаживаются изменения в переднедорсальном гиппокампе и ретикулярной формации среднего мозга.
Так как в стадии концентрации оборонительных и пищевых условных рефлексов изменения электрических потенциалов в ретикулярной формации ствола мозга обычно становятся слабо выраженными, можно предположить (Л. Г. Трофимов и сотр., 1958, 1959; Т. С. Наумова, 1958, 1960; John, Кillam, 1959; Hears и сотр., 1960), что роль в замыкании неспецифических структур на этой стадии концентрации условных рефлексов постепенно ими теряется. Напротив, в системе замыкающихся анализаторов и особенно в системе анализатора, воспринимающего условный сигнал, выявляются изменения электрических потенциалов, явно свидетельствующие об активной роли их в замыкании. Эти наблюдения явились электрографической иллюстрацией положения школы И. П. Павлова, что в замыкании условного рефлекса главную роль играют анализаторы, вступающие во временную связь. Подтвердили этот взгляд и более поздние эксперименты, в которых исследовались влияния раздражения и разрушения различных подкорковых образований мозга на те или иные условные рефлексы. Разбор этих экспериментов дан в нашем обзоре "Физиология ретикулярной формации" (1963).
Как выяснилось в наших наблюдениях, преимущественная активация колебаний электрических потенциалов в корковом конце кожно-двигательного анализатора и системе слухового анализатора возникает сначала на следах очередных сочетаний, а затем совпадает с действием сигнального раздражения. Следовые процессы особенно отчетливо выступают на корковом (при записях погружными электродами) и на подкорково-стволовом уровнях сигнального анализатора. Иными словами, если на первых двух стадиях формирования условного рефлекса (стадиях генерализации) следовые реакции (сначала депрессия, а затем экзальтация) носили диффузный характер, то в III стадии (стадии концентрации) они приурочиваются преимущественно к анализаторам, вступающим во временную связь.
В ту доминантную констелляцию, которая формируется в III стадии (первом периоде концентрации) условного рефлекса, включается целая система структур мозга, а не один только корковый конец двигательного анализатора, как можно было бы думать, исходя из предположений, что в основе замыкательного процесса лежит принцип моторной доминанты, или не только корковая территория сигнального анализатора, как можно было бы думать, исходя из предположения, что в основе замыкательного процесса лежит принцип сенсорной доминанты. Электрофизиологические исследования показали, что в стадии концентрации рефлекса происходит преимущественная активация электрических процессов в корковых компонентах сигнального и безусловнорефлекторного анализаторов как при осуществлении условных реакций, так и в межсигнальные периоды (М. Н. Ливанов, К. Л. Поляков, 1945; М. Н. Ливанов и сотр., 1951; М. Н. Ливанов, 1958, 1960; Р. М. Мещерский, 1955; М. Я. Рабинович, Л. Г. Трофимов, 1957). Последнее дало основание М. Я. Рабиновичу и Л. Г. Трофимову говорить, что доминантными свойствами обладает вся констелляция нервных центров, включающая в первую очередь кортикальные компоненты взаимодействующих анализаторов.
Разберем более подробно те процессы, которые возникают на уровне слухового анализатора, на уровне кожно-двигательного анализатора, а также на уровне прочих исследованных формаций мозга. Что касается слуховой системы, то, по нашим данным, активация процессов, наступающая в ответ на действие условного сигнала, а также в межсигнальные периоды, охватывает в стадии концентрации рефлекса не только кортикальные уровни взаимодействующих анализаторов, но также подкорковые и стволовые структуры сигнального (слухового) анализатора - кохлеарные ядра, трапециевидное тело, ядра верхнего оливарного комплекса, ядра латеральной петли, внутренние коленчатые тела. Эти наблюдения дают возможность утверждать, что в формирующуюся доминантную констелляцию входят не только корковые отделы анализаторов, вступающих во временную связь, но также субкортикально-стволовые звенья слуховой (сигнальной) системы. Процесс последействия в слуховом анализаторе оказался особенно выраженным. Активация слухового анализатора, наступающая одновременно во всех его звеньях, может длиться от секунд до десятков часов.
Появление длительных следовых реакций в слуховом анализаторе в процессе формирования оборонительного условного рефлекса свидетельствует о том, что в первую очередь все звенья сигнального анализатора являются носителями того следового возбуждения, которое возникает вследствие взаимодействия в нем процессов слухового и болевого происхождения. Избирательная активация слухового анализатора, наступающая как при осуществлении условной реакции на звук, так и после сочетаний определяется, по-видимому, изменениями функциональных свойств сигнального анализатора. Нам представляется, что длительная следовая активация электрических процессов анализатора условного сигнала свидетельствует о том, что этот процесс связан с собственно условнорефлекторными механизмами, ибо при частом изолированном применении индифферентного звукового раздражителя хотя и удается иногда наблюдать избирательное повышение уровня электрической активности в слуховом анализаторе, однако такое усиление активности имеет место в пределах одного опытного дня.
На каких уровнях сигнального анализатора происходит настройка к восприятию условного раздражения? Судя по экспериментам А. А. Гюрджан и других, уже в рецепторе восприятие условного сигнала происходит по-иному, нежели восприятие того же раздражителя, но не являющегося сигналом той или иной реакции. Исходя из ряда электрофизиологических наблюдений (Gastaut и сотр., 1956; Hernandez-Peon и сотр., 1956а, б; наши данные), в которых обнаружились различия между изменениями электрической активности кохлеарных ядер при действии индифферентного звука и того же звука, но ставшего сигналом условной оборонительной реакции, можно предположить, что изменения чувствительности происходят, по-видимому, и на уровне первичных переключательных реле сигнального анализатора. И, наконец, данные о зависимости электрических процессов кортикального уровня сигнального анализатора от того, является ли данный раздражитель индифферентным или сигналом условной реакции (М. Н. Ливанов, К. Л. Поляков, 1945; М. Н. Ливанов и сотр., 1951; В. В. Артемьев, Н. И. Безладнова, 1952; О. В. Верзилова, 1955; Р. Н. Лурье и сотр., 1956; А. И. Ройтбак, 1958; А. Б. Коган, 1960, и др.), позволяют думать, что процессы настройки касаются и этих отделов мозга.
Наблюдавшееся в наших опытах иногда многочасовое возрастание электрической активности в системе слухового анализатора дает основание утверждать, что изменение функционального состояния сигнального анализатора определяется длительным сохранением следового возбуждения на всех его уровнях: в кохлеарных ядрах, трапециевидном теле, верхнем оливарном комплексе, ядрах латеральной петли, заднем двухолмии, внутренних коленчатых телах и коре сильвиевой и эктосильвиевой извилин. Следовое возбуждение, являясь подпороговым, создает состояние, при котором анализатор реагирует на сигнальные раздражения по-иному, чем до выработки на них условного рефлекса. В отношении вызванных первичных ответов это выражается в повышении их величины и длительности на первые щелчки в серии, представляющей сигнал оборонительной реакции.
Возрастание вызванных первичных ответов происходит во второй и третьей фазе формирования оборонительной условной реакции (см. рис. 28). Если фазе генерализованной экзальтации электрических потенциалов (II стадия образования рефлекса) соответствовал эффект увеличения вызванных первичных ответов, то в III стадии (первом периоде концентрации) вызванные ответы возрастали еще в большей степени. Возрастание величины и длительности вызванных потенциалов в начале действия условного сигнала свидетельствует о закономерной связи их с процессом условнорефлекторного возбуждения. Особая выраженность этих ответов во второй и третьей фазах формирования условной реакции говорит о том, что активный анализ сигнала условной реакции совпадает именно с этими стадиями выработки рефлекса, когда еще не установился высокий процент правильных условнорефлекторных ответов. Об этом же свидетельствует то, что величина латентного периода появления вызванных первичных потенциалов в сигнальном (слуховом) анализаторе остается уменьшенной по сравнению с ее значениями до выработки рефлекса. Очевидно, и в этой стадии возбудимость сигнального анализатора продолжает оставаться повышенной. Но в связи с тем, что к концу III стадии наступает восстановление латентного периода первичных ответов до исходной величины, можно думать, что к концу этого периода происходит постепенное восстановление возбудимости сигнального анализатора до исходных значений.
Каково же происхождение и механизм условнорефлекторной настройки сигнального анализатора? Как было показано еще школой И. П. Павлова, замыкание условного рефлекса происходит вследствие встречной иррадиации возбуждения из очага условного и безусловного раздражений. Вследствие длительно и постоянно повторяющихся сочетаний стимулов между корковыми концами взаимодействующих анализаторов устанавливается двусторонняя связь (И. П. Павлов; И. С. Бериташвили, 1922, 1947, 1960; П. С. Купалов, 1947, 1955, 1960; Э. А. Асратян, 1959; М. Е. Варга, 1955; Е. И. Плонская, 1958; Е. Н. Соколов, 1958, 1959; А. Б. Коган, 1960; А. И. Ройтбак, 1960; Г. В. Скипин, 1960, и др.)
Долгое время механизм и значение обратных связей, формирующихся в направлении от коркового очага безусловного к корковому очагу условного раздражения, не подвергались анализу. Лишь в последнее время обратным связям начали придавать известное значение в формировании поступательных условных рефлексов. Так, П. С. Купалов (1955), Э. А. Асратян (1959), Е. Н. Соколов (1959), А. Б. Коган (1960) высказали взгляд, что условнорефлекторные изменения в процессах коркового конца сигнального анализатора происходят под влиянием обратных связей.
Нельзя не согласиться с такой трактовкой значения связей обратного направления. Однако нам представляется, что значение их не ограничивается только влиянием на корковый конец анализатора, воспринимающего условный сигнал. По-видимому, эти связи играют определенную роль в настройке всего анализатора к восприятию условного сигнала. Как представить себе механизм этого явления? Остановимся, например, на процессах, происходящих в нижних звеньях слухового анализатора и, в частности, в кохлеарных ядрах.
Исходя из факта условнорефлекторного изменения уровня их возбуждения, а также длительного сохранения этого возбуждения, можно полагать, что уже на уровне первичного переключательного реле слухового анализатора происходит взаимодействие афферентаций, вызванных с разных анализаторов. Прямых наблюдений конвергенции импульсов или электротонических влияний разных модальностей в кохлеарных ядрах пока что не известно. Но по аналогии с ядрами клиновидного канатика можно полагать, что конвергенция импульсов возможна и на уровне кохлеарных ядер. Известно, что кохлеарные ядра, как и другие специфические образования, получают коллатерали аксонов клеток ретикулярной формации (В. М. Бехтерев, 1896; Ramon у Cahal, 1909; М. Е. Шайбель, А. Б. Шайбель, 1958), через которые может поступать в эти образования неспецифическая для них информация. Показано также влияние ретикулярной формации на восприятие и проведение возбуждения в специфических системах и, в частности, кохлеарных ядрах. Так, Galambos и сотрудники, Hernandez-Peon и сотрудники обратили внимание на изменения вызванных потенциалов первичных слуховых реле (кохлеарных ядрах) в зависимости от фона наличной афферентации (реакции внимания). Они показали, что угнетение афферентного потока в сенсорных путях ствола при действии внешних раздражений зависит от ретикулярных влияний (Hernandez-Peon и сотр., 1956а; Galambos и сотр., 1956; Brust-Carmona, Hernandez-Peon, 1959), так как раздражение ретикулярной формации угнетает вызванные потенциалы кохлеарных ядер и других первичных сенсорных реле. По данным Galambos (1956), это влияние опосредуется tr. olivocochlearis. Вопрос о точках приложения ретикулярных влияний на проведение возбуждения в основных афферентных путях до настоящего времени не решен. Так, Hugelin и сотр. (1959, 1960) показали, что уменьшение вызванных потенциалов не наблюдается, если удалены мышцы среднего уха. Эти данные позволили заключить, что ретикулярная формация не меняет характера слуховой информации в подкорково-стволовых инстанциях, но влияет на восприятие в рецепторе. Так, по мнению большинства исследователей, ретикулярная формация оказывает преимущественно тормозящее влияние на восприятие в рецепторе и (или) в первичных реле анализаторов. Исходя из этих данных трудно было бы думать, что возбуждение, исходящее из ретикулярной формации, способно оказать на кохлеарные ядра влияние, сенсибилизирующее их к действию условного сигнала. С другой стороны, имеется мнение, что нисходящее влияние ретикулярной формации может быть активирующим. Так, А. М. Марусева (1961) отметила, что снижение амплитуды вызванного потенциала кохлеарных ядер на фоне ориентировочной реакции сопровождается укорочением скрытого периода ответа и времени восстановления возбудимости, свидетельствующее, по мнению автора, о нисходящем активирующем влиянии ретикулярной формации. Такие наблюдения немногочисленны и только в том случае, если они подтвердятся, можно будет думать и об облегчающем нисходящем влиянии ретикулярной формации на восприятие, а пока что значительно больше фактов, говорящих о тормозящем нисходящем влиянии этого образования на проведение возбуждения в нижних звеньях сигнального анализатора и, в частности, в кохлеарных ядрах.
Таким образом, ретикулярная формация не может являться той системой, которая формирует условнорефлекторную настройку нижних отделов слухового анализатора.
Вместе с тем, взаимодействие звуковой и болевой афферентации в кохлеарных ядрах может опосредоваться ретикулярной формацией в том смысле, что болевая импульсация, проводящие пути которой идут главным образом в пределах ретикулярной формации ствола мозга (С. Дж. Херрик, Г. Г. Бишоп, 1958), имеет большие возможности для непосредственного влияния на первично переключательные аппараты и другие стволовые отделы слухового анализатора по принципу обратных связей, т. е. связей, идущих из системы кожно-двигательного к слуховому анализатору, осуществляющихся и на подкорково-стволовом уровне. О том, на какие элементы субкортикальных отделов слухового анализатора преимущественно влияет болевая афферентация, сказать без специальных исследований трудно, но не исключена возможность, что не последнюю роль играют в этом отношении ретикулярные нейроны, которые встречаются в самых различных звеньях специфических систем (Г. П. Жукова, 1959; Т. А. Леонтович, 1959; Е. Г. Школьник-Яррос, 1959).
Однако возможен и другой механизм, который сводится к ретроградному влиянию на кохлеарные ядра и другие звенья слуховой системы возбуждения, синтезированного на высших этажах нервной системы. Откуда эти влияния могут осуществляться? Теоретически со всех более высоких уровней мозга, но наибольшее значение в этом смысле имеют, по-видимому, высшие отделы слухового анализатора, осуществляющие свои влияния с помощью кортикофугальных, геникулофугальных, бигеминофугальных путей. Особенно в этом смысле значимы кортикофугальные влияния. Как известно из работ школы И. П. Павлова, корковые структуры оказывают существенное влияние на деятельность стволово-подкорковых отделов мозга. Наблюдения последних лет показали, что это в одинаковой мере касается как неспецифических (Дж. Д. Френч, 1958; Р. Б. Ливингстон, 1958; С. П. Нарикашвили, 1960), так и специфических (Дж. Вада, 1958; Ogden, 1960) систем. Хотя Дж. Вада (1958) и утверждает, что в ретикулярной формации больше клеток, реагирующих на кортикальную стимуляцию, чем в специфических ядрах (nucl. ventralis posterolateralis), иными словами, неспецифические системы в большей мере находятся под контролем коры, чем специфические, тем не менее специфические системы также находятся под ее определяющим воздействием. Полагают (Ogden, 1960), что кора головного мозга контролирует проведение возбуждения в специфических системах по-иному, чем ретикулярная формация. Данные о кортикофугальных влияниях на проведение "специфического" возбуждения получены главным образом в отношении зрительной системы (Widen, Ajmone-Marsan, 1961; Р. М. Мещерский и сотр., 1963). Показано, что эти влияния являются как облегчающими, так и тормозящими. Нисходящие влияния коры на субкортикальные отделы слухового анализатора также могут быть и облегчающими и тормозящими (Desmedt, Mechelse, 1958, 1959).
Ведущая роль в отношении влияния на слуховую систему, очевидно, принадлежит слуховой зоне коры головного мозга. Это заключение исходит из того наблюдения, что эфферентные пути каждого анализатора берут начало преимущественно (но не исключительно) от территорий, принадлежащих к данному анализатору. Об этом же говорят факты значительного нарушения условнорефлекторной деятельности и особенно дифференцировочного торможения в результате всякого рода повреждений слуховой коры (М. Э. Эльясон, 1908; И. С. Маковский, 1908; И. И. Крыжановский, 1909; Kryter, Ades, 1943; Raab, Ades, 1946; Tunturi, 1955; Butler и сотр., 1957; Neff, 1958; T. A. Meринг, 1960). Особенно страдают только что выработанные условные рефлексы (Stepien и сотр., 1960). Однако эти же эксперименты показали, что осуществление условных рефлексов у животных, лишенных коркового конца сигнального (слухового) анализатора, все-таки возможно. Это свидетельствует о том, что в случае поломки коркового конца саморегуляция и анализ наиболее тонких сигналов исчезают, однако сохраняются более грубые процессы настройки. Способность организма вступать во временные функциональные взаимоотношения со средой на более грубые ее сигналы сохраняется вследствие запасных, дублирующих устройств на более низких уровнях анализатора, в которых процессы настройки на восприятие условного сигнала осуществляются также вследствие взаимодействия в них влияний слухового и болевого происхождения (в случае оборонительной реакции).
Итак, условнорефлекторные процессы настройки сигнального анализатора на восприятие условного звука происходят вследствие взаимодействия (замыкания) в нем нервных процессов, вызываемых сочетанием звукового и болевого раздражений. На возможность установления временной связи между элементами в пределах одного анализатора указывали О. С. Адрианов (1952); Н. Н. Дзидзишвили (1956, 1960), Е. Н. Соколов (1960). Нам представляется, что эта связь в системе сигнального анализатора формируется под определяющим влиянием со стороны безусловнорефлекторного анализатора. По-видимому, замыкание в пределах одного (в частности, сигнального) анализатора является закономерным процессом при формировании временной связи.
На всех стадиях выработки рефлекса обращают на себя внимание сопряженные изменения электрической активности разных звеньев слуховой системы, наступающие как в ответ на действие условных раздражений, так и в последействии сочетаний. Эти наблюдения говорят о том, что слуховой анализатор претерпевает условнорефлекторные изменения на всем своем протяжении от кохлеарных ядер до слуховой коры. Чем определяется такая общность реакций? Определяется ли она тем, что звуковой сигнал встречает на всем протяжении слухового анализатора состояние повышенной реактивности, обусловленное взаимодействием процессов в каждом звене слуховой системы, или состояние повышенной восприимчивости всего анализатора определяется повышением возбудимости только его рецепторного отдела, либо, наконец, сенсибилизация анализатора происходит под влиянием возбуждения, синтезированного на более высоких уровнях, и особенно корковом? Ответить на все эти вопросы без специальных экспериментов не представляется возможным. Однако наши наблюдения дают основание думать, что все эти процессы имеют место в известной временной последовательности, ибо каждое воздействие сигнального раздражения сначала вызывает общее изменение электрической активности - генерализованную депрессию или экзальтацию (в зависимости от индивидуальных особенностей, функционального состояния нервной системы животного и интенсивности сочетаемых раздражений), которые можно сопоставить с возникновением ориентировочной реакции (Е. Н. Соколов, 1958), ведущей к повышению возбудимости всех анализаторов, или распространением опережающего возбуждения (П. К. Анохин, 1962). Эти сдвиги, как уже отмечалось, определяются, по-видимому, взаимодействием процессов на всех, начиная с самых низших, уровнях анализатора. Локальные условнорефлекторные изменения электрической активности сигнального анализатора наступают лишь как следствие этой предварительной генерализованной волны возбуждения, распространяющейся, как уже говорилось, с нижних отделов мозга в направлении к коре. Специфические условнорефлекторные сдвиги в сигнальном анализаторе, т. е. появление в ответ на звуковое условное раздражение эффекта локальной его активации, происходят, таким образом, позднее. Это дает основание полагать, что избирательная реакция всей слуховой системы наступает лишь после того, как осуществился анализ поступившего звукового сигнала. Такой анализ, т. е. сопоставление с предварительно сложившейся моделью возбуждения, вызванной сочетанием двух раздражений (П. К. Анохин, 1949; Е. К. Соколов, 1959), по-видимому, осуществляется кортикальным уровнем анализатора, хотя одновременно тот же процесс происходит и в его субкортикально-стволовых звеньях.
Как следует из концепции П. А. Анохина (1958а), при выработке условной реакции создаются такие отношения, при которых с каждым новым применением условного раздражения вместе с эффекторными аппаратами условной реакции одновременно приводится в возбуждение добавочный афферентный аппарат, который точно воспроизводит качественные особенности всегда применявшегося подкрепления. Этот корковый афферентный аппарат условного возбуждения оказывается чем-то вроде контрольного аппарата, ибо в момент прихода обратных афферентных импульсаций от безусловного раздражения в кору он оценивает степень совпадения между этим добавочным афферентным комплексом условного рефлекса и подкреплением. Этот аппарат, производящий оценку корой головного мозга любого рефлекторного акта, был назван П. К. Анохиным "акцептором действия". Наши наблюдения дают основания полагать, что в широком смысле слова акцептором действия является вся система структур, участвующая в формировании условного рефлекса, и в понятие акцептора действия, таким образом, нельзя не включить систему условного анализатора, деятельность которого находится под непосредственным контролем безусловной, подкрепляющей афферентации. По-видимому, и в этом смысле главная роль принадлежит его корковому концу. Лишь в результате совпадения сигнала с ранее сформировавшейся моделью возбуждения происходит активация анализатора условного сигнала. Импульсация анализатора, воспринимающего условный сигнал, усиливается и благодаря поступлению дополнительной афферентации со смежных анализаторов. Об этих механизмах уже шла речь в предыдущей главе. Активация сигнального анализатора сказывается на параметрах вызванных ответов.
В III стадии формирования рефлекса, так же как и в I и II стадиях, в момент осуществления условной эффекторной реакции происходит уменьшение величины, длительности и латентного периода вызванных ответов в первичной корковой проекционной зоне сигнального (слухового) анализатора (см. рис. 28, ответ на 3-й щелчок), свидетельствующее о повышении скорости возникновения и проведения условнорефлекторного возбуждения в течение действия условного сигнала. В предыдущей главе уже говорилось о том, что этот процесс определяется повышением возбудимости и лабильности сигнального анализатора в процессе подготовки и осуществления условной реакции, а также, возможно, включением в проведение условнорефлекторного возбуждения путей с меньшим количеством переключений по их ходу.
Условнорефлекторной настройки сигнального анализатора, однако, недостаточно ни для осуществления условной реакции, ни для полной оценки текущего рефлекторного акта. Процесс активации слухового анализатора приводит к усилению иррадиации возбуждения в направлении к кожно-двигательному анализатору, что тем самым обеспечивает вовлечение последнего в доминантный процесс. В последнем также происходит взаимодействие процессов возбуждения, вызванных условным и безусловным раздражениями в системе кожно-двигательного анализатора. Взаимодействие афферентаций со слухового и кожно-двигательного анализаторов возможно теоретически на всех уровнях последнего, т. е. в системе восприятия и проведения болевой и тактильной афферентации.
Однако в наших экспериментах было обнаружено, что ни одно из субкортикальных звеньев кожно-двигательного анализатора не входило в доминантную констелляцию, которая складывалась на III этапе образования рефлекса. В нее включались лишь корковые уровни этого анализатора.
Как показали результаты декортикации и экстирпации сенсомоторной зоны коры, осуществление локальных оборонительных условных рефлексов сильно страдает после таких вмешательств (В. П. Протопопов, 1909, 1931; Н. И. Афанасьев, 1913; Д. С. Фурсиков и М. Н. Юрман, 1925а, б; Д. С. Фурсиков, 1925; А. И. Шумилина, 1949а, б, в; О. Загер и сотр., 1961; М. И. Самойлов, 1962; А. С. Батуев, 1967). Хотя есть наблюдения, которые говорят о том, что локальные реакции после разрушения коркового конца кожно-двигательного анализатора могут восстанавливаться (Э. А. Асратян, 1934; И. С. Розенталь, 1941; К. С. Абуладзе, И. С. Розенталь, 1948; В. М. Касьянов, 1955; Л. С. Гамбарян, 1959; А. С. Батуев, 1967, и др.), их длительное выпадение свидетельствует о значении этой корковой зоны для формирования условного оборонительного рефлекса. Приведенные выше данные (одни из которых послужили основанием, а другие подтвердили выводы школы И. П. Павлова) показали, что процессы взаимодействия звуковой и тактильно-болевой афферентации, определяющие замыкание оборонительного условного рефлекса, происходят в кортикальных компонентах кожно-двигательного анализатора. Электрофизиологические наблюдения подтверждают эти заключения. Предполагают, что в соматосенсорной зоне коры тесно связаны и рецепторные элементы разных анализаторов и эфферентные элементы кожно-двигательного анализатора, что создает благоприятные условия для конвергенции процессов на ее уровне (Albe-Fessard, 1957; Бюзер, Эмбер, 1961; А. С. Батуев, 1967), ибо соматосенсорная афферентация достигает проекционных зон (SI, SII, SIA), которые представляют области перекрытия с моторной коры (Albe-Fessard, 1957). Ничто не отличает зоны SI, SII, SIA в их способности отвечать на стимуляцию кожного и мышечного происхождения (Bremer, 1952; Mountcastle и сотр., 1952). В зоне SII оканчивается и путь болевой афферентации (Biemond, 1956), правда, по данным Д. М. Гедеванишвили (1959), его окончание следует отнести и к зрительной области коры головного мозга.
Наши наблюдения согласуются с ранее полученными данными (М. Н. Ливанов, К. Л. Поляков, 1945; М. Н. Ливанов и сотр., 1951; В. В. Артемьев и Н. И. Безладнова, 1952; Р. Н. Лурье и сотр., 1956; О. В. Верзилова, 1958, и др.), что в сенсомоторной зоне коры при образовании условного оборонительного рефлекса происходят изменения электрической активности (появление вызванных ответов и активация колебаний потенциала), свидетельствующие о несомненном участии этой структуры в образовании рефлекса.
Что касается участия кожной и проприоцептивной афферентации в формировании доминантной констелляции в стадии концентрации, то мнения исследователей по этому вопросу довольно противоречивы. И. М. Аптер (1927), В. П. Протопопов (1931), Л. С. Гамбарян (1959) считают, что проприоцептивная импульсация играет большую роль в формировании условного двигательного рефлекса. Однако, по наблюдениям А. А. Ющенко (1933), Н. В. Раева и Е. Я. Раппопорта (1934), разрушение задних столбов не приводит к выпадению условных двигательных реакций. По мнению этих авторов, а также Я. Буреша и О. Бурешовой (1960), это объясняется тем, что проприоцептивная импульсация не играет важной роли в формировании двигательных рефлексов. Л. С. Гамбарян (1959) отстаивает противоположную точку зрения, утверждая, что проприоцептивная сигнализация может осуществляться как через систему задних столбов спинного мозга, так и через другие спинальные пути и, в частности, рассеянные элементы проприоцептивной сигнализации в боковых и передних канатиках спинного мозга, поэтому эксперименты с разрушением задних столбиков не вызывают изменений условнорефлекторных реакций. Тем не менее, наблюдения А. А. Ющенко (1933), Н. В. Раева и Е. Я. Раппопорта (1934) приводят к выводу, что основные афферентные пути кожно-двигательного анализатора не включаются органически в систему условнорефлекторной дуги. Более того, наблюдения Э. А. Асратяна (1935) показали, что в момент осуществления условной оборонительной реакции возбудимость кожно-двигательного анализатора падает, ибо порог оборонительной реакции на фоне условнорефлекторного влияния звука повышается. Эти факты дали возможность Э. А. Асратяну (1937) высказать мысль, что... "афферентная часть дуги безусловного рефлекса не является абсолютно необходимой для существования и проявления условного рефлекса". Нам пришлось убедиться в справедливости этого предположения применительно к III стадии оборонительного условного рефлекса.
Как обнаружилось в наших экспериментах, в III стадии (первом периоде концентрации рефлекса) условный сигнал не вызывал существенных изменений электрической активности ни в ядрах нежного и клиновидного канатиков, ни в ретикулярных структурах среднего и продолговатого мозга, ни в центральном сером веществе, ни в вентральном ядре таламуса, т. е. ни в одном из тех образований, которые причастны к проведению соматической (кожной и проприоцептивной) (тактильной и болевой) афферентации. В этих структурах мозга, так же как в других стволовых формациях, но не принадлежащих к системе слухового анализатора, и в момент действия условного сигнала, и в период межсигнальных реакций регистрируется лишь слабый эффект активации. Эти данные дают основание утверждать, что афферентные пути кожно-двигательного анализатора не входят в ту доминантную констелляцию, которая лежит в основе оборонительной временной связи в первый период концентрации рефлекса, хотя последняя и формируется и контролируется соматосенсорной афферентацией.
Как было показано в исследовании М. Я. Рабиновича (1958), в стадии концентрации условного оборонительного рефлекса происходит преимущественная активация слоя V сенсомоторной области коры головного мозга, что свидетельствует об определяющем значении процессов именно в этом слое, богатом крупными пирамидными нейронами. Участие слоя V в создании моторной доминанты подтвердилось и в исследованиях М. А. Рябининой (1961), показавшей, что катодизацией слоя V сенсомоторной области коры мозга кролика можно вызвать создание доминантного очага.
Очевидно, активация кортикальных компонент кожно-двигательного анализатора при осуществлении оборонительного рефлекса происходит за счет процессов в эффекторных элементах сенсомоторной зоны коры, которые приводятся в доминантное состояние вследствие предшествующих многократных совпадений слухового и болевого раздражений.
Влияние слухового возбуждения на двигательный анализатор на первых трех стадиях образования рефлекса в большой мере осуществляется по внутрикорковым каналам, ибо, как показали Я. М. Прессман (1938), Н. Н. Дзидзишвили (1956), О. С. Адрианов (1960), разрез коры, прерывающий пути между корковыми территориями анализаторов, вступающих во временную связь, ведет к выпадению локальных оборонительных условных реакций. Нельзя не отметить в то же время, что есть данные, отрицающие прямые ассоциативные связи слуховой и сенсомоторной зон коры (А. С. Батуев, 1967).
Иными словами, в III стадии выработки рефлекса (первом периоде концентрации), когда уменьшается объем побочных компонент эффекторного ответа и сама условная реакция становится более локальной, на фоне сохраняющейся широкой иррадиации возбуждения приобретают определяющее значение процессы, которые протекают в анализаторах, вступающих во временную связь. В формирующуюся в этой фазе образования рефлекса доминантную констелляцию входят все уровни сигнального анализатора и эффекторные элементы коркового уровня кожно-двигательного анализатора. В III стадии выработки рефлекса следовые процессы наиболее длительно сохраняются в системе слухового анализатора, поэтому можно думать, что формирующаяся на III этапе доминантная констелляция развивается с акцентом на сенсорной доминанте в системе сигнального анализатора с преимущественным развитием обратных связей от кожно-двигательного анализатора к слуховому. Так как, по нашим данным, в сенсомоторной зоне коры процессы активации по мере развития III стадии и перехода ее в IV все более усиливаются, в то время как в системе сигнального анализатора они, наоборот, ослабляются, это свидетельствует о постепенном нарастании уровня возбуждения в эффекторной части доминантной констелляции и снижении его в афферентной ее части.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'