II стадия формирования условного оборонительного рефлекса (второй период генерализации)
По мере сочетаний условного раздражения с безусловным реакции депрессии, сопровождающей осуществление рефлекса, начинает предшествовать вспышка активности, имеющая диффузное распространение или приуроченная к системе взаимодействующих анализаторов (слуховому и двигательному). При дальнейшей тренировке условного рефлекса экзальтация активности все более и более замещает собой реакцию угнетения, и усиление электрических колебаний наступает во всех исследованных подкорково-стволовых ядрах и территориях коры, регистрируемых с помощью погружных электродов. I и II стадии выработки рефлекса переходят друг в друга постепенно, так что, кроме "чистых" случаев, можно видеть картины, несущие в себе черты I и II этапа.
Как и в I стадии, наблюдается сходство изменений ЭГ в стволовых отделах слухового, двигательного анализаторов и ретикулярной формации каждого уровня мозга.
Фаза генерализованной экзальтации активности при выработке рефлексов в ответ на звуковые раздражения пороговой интенсивности обнаружена у всех подопытных животных. Из этих собак у 6 II стадия наступила при первых же сочетаниях раздражителей (см. табл. V).
Описание изменений ЭГ структур мозга на этой стадии выработки рефлекса мы начнем с тех животных, у которых эта реакция пришла на смену реакции генерализованной депрессии электрической активности. Так, у собаки № 4 105-е применение порогового звукового сигнала (рис. 25) сопровождается отчетливо выраженным возрастанием электрической активности в ядрах нежного и клиновидного канатиков, кохлеарных ядрах, нижнем центральном ядре ретикулярной формации, коре сигмовидной, эктосильвиевой, супрасильвиевой и латеральной извилин больших полушарий головного мозга. Возрастание активности в эктосильвиевой и супрасильвиевой извилинах выражается главным образом в повышении частоты электрических осцилляций (с 48-50 до 55-60 в 1 сек). Следует обратить внимание на значительное сходство изменений ЭГ ядер нежного и клиновидного канатиков, кохлеарных ядер и нижнего центрального ядра ретикулярной формации. Нельзя не отметить также, как близки по характеру изменения ЭЭГ сенсомоторной зоны коры, регистрируемые погружными и накладными электродами, чего не наблюдалось на предыдущей стадии выработки рефлекса (рис. 14). Направление изменений ЭЭГ в сенсомоторной зоне коры левого и правого полушарий общее, однако степень возрастания активности в левой двигательной области, по крайней мере, в 2 раза выше, чем в правой. Периоду генерализованного роста электрических осцилляций предшествует длящийся 100-150 мсек период их угнетения. Первые проявления тонической активности мышц правой передней лапы совпадают с началом генерализованного усиления колебаний. Эффект активации сохраняется несколько секунд и впоследействии раздражения. Фаза генерализованной экзальтации активности у собаки № 4 была крайне непродолжительной (105-107-е сочетание), однако ее появление отмечалось и на более ранних этапах выработки рефлекса (см. рис. 15, а, б).
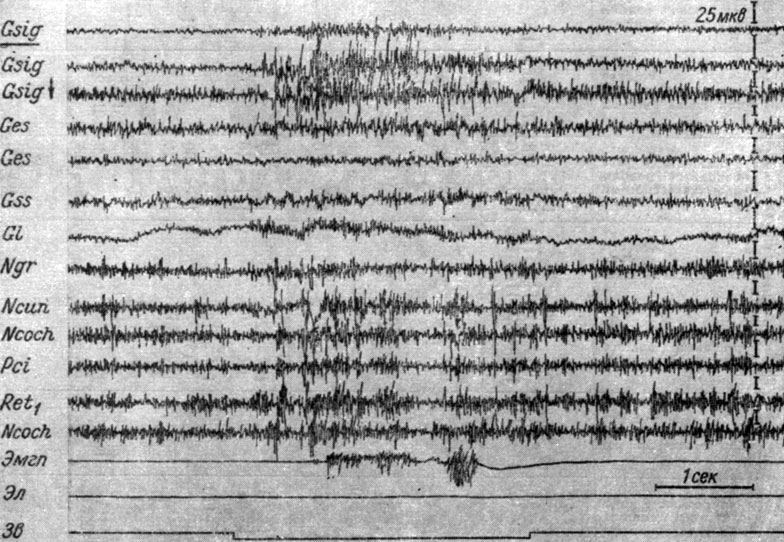
Рис. 25. Изменения ЭГ разных образований коры большого и продолговатого мозга собаки № 4, наступающие во II стадии формирования условного оборонительного рефлекса. 105-е применение тона 500 гц 48 дб (опыт № 35 от 27/II 1959 г.)
Сопоставление описываемых изменений электрической активности разных структур мозга с вызванными ответами в первичной проекционной корковой зоне слуха показало, что во второй фазе выработки рефлекса наступает возрастание величины вызванных потенциалов. Сказанное иллюстрирует рис. 16, на котором видно, что у собаки № 4 увеличение вызванных ответов в первичной слуховой зоне коры на включение условного стимула (тона 500 гц 48 дб) происходит к 107-110-му сочетаниям, когда и по показателям изменений основной электрической активности происходит переход к фазе генерализованной экзальтации электрических процессов.
Длительность следовых процессов во второй стадии выработки рефлексов не превышает нескольких секунд (см. рис. 18).
Как отмечалось, II фаза генерализации в ряде случаев наступала почти с самого начала сочетаний звукового сигнала с болевым подкреплением лапы. Так, у собаки № 3 уже 6-е применение условного звукового сигнала пороговой интенсивности (500 гц 48 дб) сопровождается (рис. 26, а) генерализованным возрастанием электрических колебаний (повышением их ритма и величины) в кохлеарных ядрах, ядрах нежного и клиновидного канатиков и нижнем центральном ядре ретикулярной формации, которому предшествует четверть секундный период их слабого снижения. Весь период действия условного сигнала в кортикальных отведениях накладными электродами регистрируется повышение ритма осцилляций при одновременном незначительном снижении их амплитуд. Различия в характере реакции корковых и стволовых структур объясняется различиями в способе регистрации их электрической активности. Интересное явление наблюдается в ЭЭГ коры латеральной извилины. В ней появляются двухфазные отклонения (в верхней части ограниченные движением пера), сходные с теми, которые возникают в указанных отделах мозга при световом раздражении.
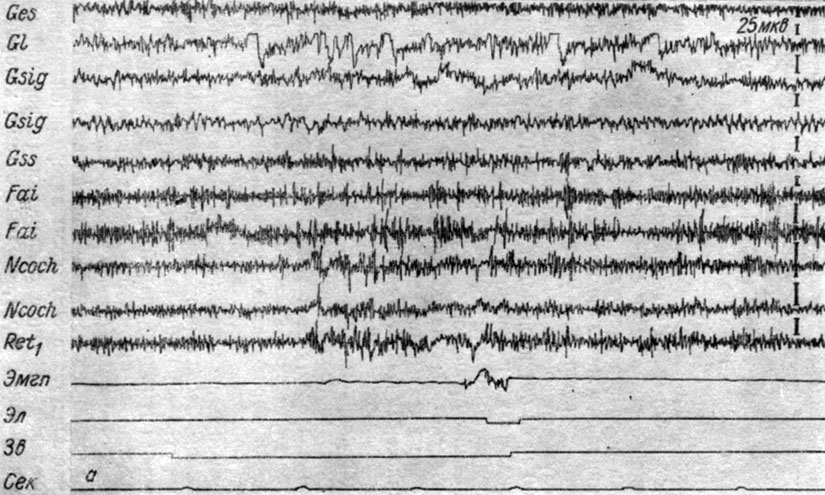
Рис. 26. Изменения ЭГ коры большого мозга и структур продолговатого мозга собаки № 3, наступающие во II (а) и III (б) стадиях формирования условного оборонительного рефлекса. а - 6-е сочетание тона 500 гц 48 дб с электрокожным раздражением правой передней лапы (опыт № 5 от 23/1 1959 г.); б - 83-е сочетание тех же раздражений (опыт № 27 от 27/II 1959 г.) (по Т. С. Наумовой, 1962в)
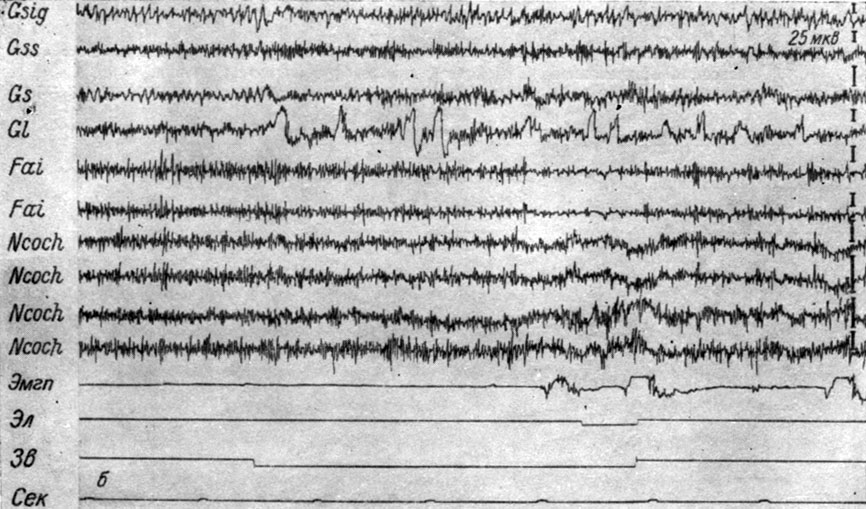
Рис. 26. Продолжение
Вызванные потенциалы в первичной проекционной зоне слуха (коре эктосильвиевой извилины), а также сенсомоторной зоне (коре сигмовидной извилины), появляющиеся на включение условного сигнала (тона 500 гц 48 дб) во II стадии выработки условного рефлекса, несколько возрастают по сравнению с ответами в I стадии (рис. 19) и у собаки № 3.
Эффект генерализованной экзальтации активности во II стадии формирования рефлекса имеет место и на других уровнях мозга. На рис. 27 представлены ЭГ корковых и мезенцефалических структур собаки № 2, зарегистрированные при 20-м сочетании условного звукового сигнала пороговой интенсивности (500 гц 48 дб) с пороговым электрокожным раздражением. Видно, что действие условного стимула вызывает возрастание электрической активности, в одинаковой степени выраженное в заднем двухолмии, внутреннем коленчатом теле, центральном сером веществе и ретикулярной формации среднего мозга. Отмечается нарастание амплитуд и частот колебаний и в коре коронарной и супрасильвиевой извилин.
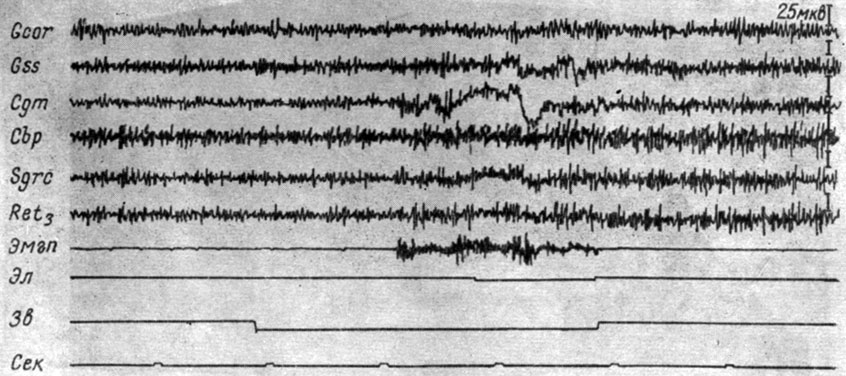
Рис 27. Изменения ЭГ коры большого мозга и структур среднего мозга собаки № 2, наступающие во II стадии образования условного оборонительного рефлекса 20-е сочетание тона 500 гц 48 дб с электрокожным раздражением правой передней лапы (опыт № 49 от 10/XII 1958 г.)
Как и на включение условных сигналов, вызванные ответы на ритмическое их действие в первичной проекционной зоне слуха испытывают отчетливые изменения по сравнению с I периодом формирования условной реакции.
Анализ параметров (амплитуды, латентного периода и длительности) их первых трех компонент в первичной проекционной зоне слуха на условные ритмические (2 в 1 сек) звуковые раздражения (щелчки) у собак показал, что во второй фазе образования условного оборонительного рефлекса происходят их значительные изменения на все 6 щелчков в серии, служащей условным сигналом. Что касается трех первых компонент ответов на щелчки в период изолированного действия раздражения, то их величина во II периоде формирования условного рефлекса значительно возрастает. Оказанное иллюстрируют рис. 21, в, а также рис. 22 (собака № 19). Как следует из рисунков, во II стадии образования рефлекса наблюдается увеличение и первой позитивной, и негативной, и второй позитивной компоненты ответа на каждый щелчок в серии, служащей сигналом условной реакции. Одновременно с ростом величины происходит возрастание длительности всех компонент ответа, которая либо достигает исходной величины, либо несколько превосходит ее (см. рис. 24, длительность негативной и второй позитивной компоненты ответа). Латентные периоды начала, время до максимума позитивной фазы, а также максимума негативной фазы таких вызванных ответов, так же как в первой стадии, сохраняются уменьшенными по сравнению с фоновыми потенциалами (см. рис. 23).
Величина и длительность трех первых фаз вызванного потенциала на щелчки, предшествующие и совпадающие с осуществлением условной двигательной реакции, испытывают дополнительные изменения. По мере приближения к моменту условной реакции амплитуда и длительность всех трех компонент первичного ответа значительно уменьшается. Более подробно эти сдвиги будут охарактеризованы при описании III стадии выработки рефлекса, где они выступают более отчетливо.
Итак, условная реакция на звуковой сигнал подпороговой и пороговой интенсивности на II этапе выработки рефлекса формируется на фоне генерализованной экзальтации (возрастания частоты и амплитуд колебаний) электрической активности, обнаруживаемой в записях погружными электродами на разных уровнях мозга: кохлеарных ядрах, ядрах нежного и клиновидного канатиков, внутреннем коленчатом теле, заднем и переднем двухолмии, ретикулярной формации ствола мозга, красном ядре, черной субстанции, центральном сером веществе, вентральном и ретикулярном ядрах таламуса, субталамическом ядре, структурах обонятельного мозга (гиппокампе, миндалевидном ядре), хвостатом ядре и, наконец, в территориях коры большого мозга.
Эта форма активности появляется сначала на следах сочетаний, а затем и в ответ на действие условного звукового сигнала. Та же картина активности может сохраняться в последействии и сопровождать межсигнальные реакции. Реакция активации выступает в субкортикальных и кортикальных системах одновременно, и лишь иногда удается заметить, что в стволово-подкорковых ядрах описываемые изменения появляются на несколько десятков миллисекунд раньше, чем в коре.
Во второй период выработки рефлекса обнаруживается четкая корреляция электрических реакций в так называемых специфических и неспецифических системах мозга каждого уровня нервной системы, как-то: кохлеарных ядрах, ядрах нежного и клиновидного канатиков, нижнем центральном, мелкоклеточном и крупноклеточном ядрах ретикулярной формации продолговатого мозга или внутреннем коленчатом теле, ядрах латеральной петли, задних и передних буграх четверохолмия, ядрах ретикулярной формации среднего мозга, красном ядре, черной субстанции, центральном сером веществе и т. п.
Условнорефлекторное движение конечности может как предшествовать, так и совпадать с началом генерализованной экзальтации активности, а также появляться на фоне развития последней. Электрокожное подкрепление обычно сопровождается усилением изменений, регистрируемых при действии условного сигнала.
Генерализованное возрастание частоты и амплитуд электрических колебаний сменяет фазу генерализованной депрессии активности (если таковая у животного имеется) лишь в случае относительно слабых звуковых сигналов (подпороговой или пороговой интенсивности). Если условные раздражения по интенсивности значительно превышают пороговые значения, а также при сильных болевых подкреплениях, стадии генерализованной экзальтации активности не проявляется, то первая электрографическая стадия непосредственно сменяется третьей.
Анализ параметров вызванных ответов в первичной проекционной зоне слуха на включение условных звуковых раздражений или серию щелчков показал, что во II стадии формирования условного оборонительного рефлекса происходит увеличение (по сравнению с изменениями на I стадии) амплитуды и длительности первых трех компонент (позитивной, негативной и второй позитивной) вызванных ответов. Латентные периоды таких вызванных ответов, так же как в первой стадии, сохраняются уменьшенными. Наиболее выражены эти сдвиги на включение условного сигнала (первый щелчок).
По мере приближения к моменту эффекторной условной реакции происходит редукция вызванных ответов при одновременном укорочении их длительности.
Второй период генерализации у разных собак имел разную продолжительность (см. табл. V). У 6 собак эффект экзальтации электрических колебаний в записях погружными электродами наблюдался с первых же сочетаний звукового и болевого раздражений, так что для этих животных эта стадия могла быть расценена как первая. Аналогичное явление имело место и для повторно вырабатываемых условных рефлексов на условные сигналы подпороговой интенсивности. Вторая стадия выработки рефлекса совпадает с периодом еще недостаточно устойчивых условнорефлекторных ответов.
В электроэнцефалоскопических исследованиях также показано, что в коре и подкорковых образованиях в процессе формирования условной связи происходит возрастание и увеличивается распространение местной высокоамплитудной активности (МВКП) (Г. И. Шульгина, 1960, 1961; И. Н. Книпст, 1961; М. Н. Ливанов, 1958).
Как показали наши наблюдения, условнорефлекторные изменения электрической активности в начале выработки оборонительного рефлекса регистрируются во всех исследованных пунктах подкорково-стволового и кортикального уровней мозга и в этот процесс в одинаковой степени включаются все уровни как "заинтересованных", так и "незаинтересованных" анализаторов, равно как и всех других формаций.
Нельзя согласиться, что широкая иррадиация процессов внутри мозга, которая наблюдалась в наших экспериментах и в опытах других исследователей, могла бы усиливаться, как это полагают Н. Иошии и сотрудники (Yoshii и сотр., 1957), наличием погружных электродов. Скорее можно разделить точку зрения Hagbarth и Kerr (1954), утверждающих, что бодрствующее состояние создает большие условия для иррадиации возбуждения.
В обеих стадиях генерализации нами обнаружена четкая корреляция процессов, наступающих в структурах каждого уровня мозга. Например, на уровне продолговатого мозга она была обнаружена в кохлеарных ядрах, верхнем оливарном комплексе ядер, трапециевидном теле, ядрах нежного и клиновидного канатиков, нижнем центральном, мелкоклеточном и крупноклеточном ядрах ретикулярной формации; на уровне среднего мозга - в ядрах латеральной петли, внутреннем коленчатом теле, заднем двухолмии; ядрах ретикулярной формации среднего мозга - в красном ядре, черной субстанции, nucl. cuneiformis, nucl. subcuneiformis, а также на уровне межуточного мозга - в вентральном и ретикулярном ядрах таламуса, субталамическом ядре в гипоталамусе и т. д.
Иначе говоря, как и при осуществлении ориентировочных рефлексов, при осуществлении условнорефлекторных реакций в стадиях генерализации имеет место значительное сходство электрических процессов в так называемых специфических и неспецифических системах ствола мозга, т. е. в анализаторных и, условно говоря, межанализаторных зонах мозга.
Как уже отмечалось во введении к этому разделу, большинство исследователей так или иначе наблюдали сходство изменений электрических процессов в различных субкортикальностволовых территориях мозга при действии условных сигналов в начальный период становления рефлекса (Sharpless, Jasper, 1956; Hernandez-Peon и сотр., 1956а, b, 1957; Galambos и сотр., 1956, 1960; Л. Г. Трофимов и сотр., 1958, 1959; Г. В. Мегун, 1958, и др.), но не акцентировали на нем внимание и не обсуждали его механизмы. Кроме того, в этих исследованиях не было отмечено, насколько включаются в диффузное проведение возбуждения и специфические системы "незаинтересованных" анализаторов.
В предыдущем разделе было высказано предположение и представлены доказательства того, что генерализованные изменения ЭЭГ связаны с диффузным распространением возбуждения в восходящем направлении по всему стволу мозга, т. е. не только по неспецифическим системам (как это принято считать), но также по образованиям, которые к этим системам не причисляются, и, в частности, специфическим ядрам ствола мозга, принадлежащим к "незаинтересованным" анализаторам. Все сказанное в отношении генерализованных изменений ЭЭГ, наступающих в ответ на действие безусловных раздражений, справедливо и для действия условных сигналов на первых стадиях формирования рефлекса. Если справедливо заключение, что диффузная афферентация мозга опосредуется не только неспецифическими, но и всеми другими, в том числе специфическими, системами, то можно предположить, что взаимодействие процессов осуществляется на уровне ствола не только в ретикулярной формации.
Обнаруженное нами явление конвергенции импульсов разных модальностей на уровне первичного переключательного реле кожно-двигательного анализатора - ядре клиновидного канатика - позволяет по-новому подойти к решению вопроса о встрече двух взаимодействующих возбуждений при образовании временной связи. Эти опыты, показавшие возможность конвергенции импульсов на нейронах первичного реле кожно-двигательного анализатора - ядра клиновидного канатика, свидетельствуют о том, что конвергенция импульсов, являющаяся одной из сторон взаимодействия процессов возбуждения с разных анализаторов, осуществляется и на уровне стволовых отделов специфических систем. На этом основании можно полагать, что широкая иррадиация процесса условнорефлекторного возбуждения по стволово-подкорковым и кортикальным формациям в стадии генерализации определяется взаимодействием анализаторов на уровне каждого мозгового образования и в том числе ядрах специфических систем. Условнорефлекторные изменения электрических потенциалов, по-видимому, являются результатом многократной встречи возбуждений, вызванных с сигнального и безусловного анализаторов, на уровне каждого данного образования. Этот вывод можно подкрепить наблюдениями, в которых показан факт конвергенции, т. е. встречи, импульсов разного происхождения на нейронах ретикулярной формации ствола (Amassian, De Vito, 1954; Scheibel и сотр., 1955, и др.), нейронах ядер диффузной проекционной системы таламуса, гипоталамуса, субталамуса (Gellhorn, Ballin, 1946, и др.), нейронах ядер экстрапирамидной системы (Kruger, Albe-Fessard, 1959, и др.), нейронах структур обонятельного мозга (Green, Arduini, 1954, и др.), нейронах коры (Gellhorn и сотр., 1954; Р. Юнг, 1958; Li, 1958; П. Бюзер, М. Эмбер, 1961, и др.) и, наконец, нейронах переключательных реле специфических систем анализаторов.
Так как, по-видимому, неспецифические афференты пронизывают весь ствол мозга, в том числе и основные афферентные пути в коре, в каждой структуре может осуществляться взаимовлияние процессов, вызываемых с самых различных рецепторов. Иными словами, есть все основания считать, что имеются и структурные и функциональные возможности для встречи возбуждений разного происхождения не только в коре, не только в ретикулярной формации и других ядрах неспецифических систем мозга, но практически во всех образованиях и в том числе в подкорковых ядрах взаимодействующих анализаторов. Такое "схождение" афферентаций является общим явлением в физиологии нервных образований, и речь может идти, по-видимому, не столько о качественных, сколько о количественных различиях между теми или иными формациями мозга.
Разумеется конвергенция импульсов не является единственной формой взаимодействия процессов возбуждения, вызванных с разных анализаторов, однако она свидетельствует о возможности встречи возбуждений разного происхождения на уровне элементов того или иного образования. Что касается взаимодействия неимпульсных форм активности, то данные о них известны лишь в отношении кортикальных уровней мозга (Arduini и сотр., 1957; Goldring, O'Leary, 1957; Ардуини, 1958; Н. А. Аладжалова, 1958).
Многочисленные экспериментальные факты свидетельствуют о том, что взаимодействие процессов является результатом как конвергенции импульсов на отдельных нейронах (А. Гасто, А. Роже, 1958; Fessard, 1959; А. Фессар, 1958; Hernandez-Peon, 1957; Р. Эрнандес-Пеон, 1958, и др.), так и следствием схождения электротонических влияний, выражающихся в сдвигах постоянного потенциала больших агрегатов клеточных элементов мозга (В. С. Русинов, 1953, и др.).
Генерализованные изменения ЭГ (депрессия или экзальтация) в стволово-подкорковых и кортикальных структурах наступают при действии условных раздражений сначала в нижних отделах ствола и распространяются по нему в направлении к коре со скоростью, колеблющейся в пределах десятков сантиметров в 1 сек. Так как условнорефлекторные изменения электрической активности начинаются, по нашим данным, со структур продолговатого мозга, а лишь затем (через 30-500 мсек) появляются в структурах среднего, межуточного и переднего мозга, то можно думать, что процесс возбуждения этого рода распространяется в восходящем направлении и обусловлен проведением какого-то влияния, охватывающего постепенно снизу вверх все структуры мозга. Генерализованное распространение возбуждения, согласно этим данным, происходит со скоростью, значительно различающейся для условных реакций на разных этапах их становления от 8-10 см в 1 сек до 2 м в 1 сек. С аналогичной скоростью (25-30 см в 1 сек) распространяется реакция десинхронизации в коре головного мозга человека (В. С. Русинов, 1960). В. С. Русинов рассматривает эту реакцию как выражение процесса иррадиации возбуждения.
Так как, по нашим наблюдениям, распространение диффузной волны возбуждения по мере выработки условного оборонительного рефлекса на звук охватывает мозг все в более короткие интервалы времени, это наводит на мысль о связи этого явления с образованием рефлекса. По-видимому, процессы взаимодействия афферентаций, вызываемый с анализаторов, вступающих во временную связь, осуществляются сначала на нижних уровнях нервной системы, а затем и на более высоких ее уровнях. С взаимодействием афферентаций на уровне последних связано осуществление условного рефлекса.
Что касается природы генерализованных изменений электрической активности, которые постепенно распространяются вверх по стволовым и подкорковым формациям мозга к коре, можно предположить, что они обязаны своим происхождением электротоническим сдвигам потенциала, представляющим, по мнению В. С. Русинова (1953, 1958), форму связи между нейронами, играющую главную роль при установлении временной связи. Это заключение делается на том основании, что, как полагают (Adrian, 1931; Barron, Matthews, 1938; В. С. Русинов, 1960; Caspers, 1959; Е. Н. Соколов, 1962, и др.), сдвиги постоянного потенциала лежат в основе модуляции ЭЭГ. Приведенные наблюдения подтверждают мысль В. С. Русинова, что главную роль в замыкании временных связей играют длительные электротонические процессы (или сдвиги постоянного потенциала), обусловливающие изменения фоновой электрической активности. Экспериментальное обоснование этот взгляд получил в работах В. С. Русинова и его сотрудников (Л. А. Новикова и сотр., 1952; Т. С. Наумова, 1956а, б, 1957; А. А. Соколова, 1958, 1959; Г. Д. Кузнецова и сотр., 1959; Т. Б. Швец, 1958, 1960, 1962; Р. А. Павлыгина, 1960, 1962, и др.) а также Р. С. Мнухиной (1960, 1961), Г. И. Шульгиной (1960, 1961) и др. Мысль о неимпульсной природе иррадиации возбуждения поддерживается Д. С. Воронцовым (1960), Morrell (1961) и др. Вместе с тем, генетическая взаимосвязь электротонической и импульсной активности в вопросах механизмов замыкания изучена пока еще недостаточно, и в дальнейшем она, видимо, должна получить более полное освещение.
Естественно, встает вопрос о том, зависят ли условнорефлекторные изменения ЭЭГ в каждом образовании мозга только от того, что на их уровне осуществляется взаимодействие процессов, вызванных с разных анализаторов, или еще и от того, что эти образования испытывают влияние со стороны других структур - главных участников замыкания. По-видимому, для формирования генерализованных изменений ЭЭГ немаловажное значение должно иметь и то взаимовлияние, которое осуществляется на разных уровнях мозга между специфическими и неспецифическими системами, между корковыми и подкорковыми уровнями мозга и т. д. (см. обзоры Rossi и Zanchetti, 1957; Т. С. Наумова, 1963). Очевидно, взаимовлияние структур представляет из себя второй из возможных механизмов возникновения генерализованных изменений электрической активности. Однако этот механизм вступает в действие лишь после того, как осуществляется встреча взаимодействующих возбуждений на уровне каждой структуры мозга. С другой стороны, генерализованные сдвиги ЭГ происходят вследствие того, что они выражают эффекторные процессы, реализация которых связана с деятельностью многих структур мозга.
Генерализованные изменения электрической активности поддерживаются и той "обратной" афферентацией (П. К. Анохин, 1949), которая сопутствует осуществлению любого рефлекторного акта и также диффузно охватывает мозг. Об этом свидетельствуют наблюдавшиеся в наших опытах (см. рис. 26) локальные высокоамплитудные колебания электрической активности типа вызванных ответов в зрительной зоне коры при осуществлении условных оборонительных рефлексов на звуковые раздражения. Очевидно, возникновение в зрительной зоне коры колебаний потенциалов, аналогичных ответам на световые вспышки, свидетельствует о приходе сюда возбуждения, возникающего в зрительном анализаторе. По-видимому, это происходит по следующим причинам. Безусловные и условные оборонительные рефлексы сопровождаются перераспределением тонуса мышц конечностей и тела, а затем появлением локальной двигательной реакции (П. К. Анохин, 1949; А. И. Шумилина, 1945, 1949а, б, в; М. Ф. Корякин, 1958). Одновременно с этим меняется положение головы животного и, как следствие этого, меняется освещенность его глаз, поэтому при действии условного звукового сигнала в зрительной коре возникают многократные колебания потенциалов типа вызванных ответов. Нельзя не отметить, что в других зонах мозга при осуществлении безусловных и условных оборонительных рефлексов наступает отчетливо выраженный эффект десинхронизации активности, свидетельствующий о том, что возбуждение достигает и других территорий мозга.
Таким образом, происходящие сдвиги потенциалов (типа вызванных ответов или эффект десинхронизации) в "незаинтересованных" территориях коры головного мозга свидетельствуют о том, что в смежных анализаторах не только повышается возбудимость (Е. Н. Соколов, 1958), но, как следствие этого, происходит включение их в деятельное состояние, т. е. в проведение потока, "обратной" афферентации, осуществляющейся, таким образом, не только с соматической системы (И. М. Сеченов, 1863; Н. А. Бернштейн, 1947; П. К. Анохин, 1949, 1958а), но также с других анализаторов (П. К. Анохин, 1958а). В свою очередь эти влияния повышают восприимчивость анализаторов, вступающих во временную связь, что сенсибилизирует их замыкание и корректирует их реакцию на сигналы внешней среды.
Итак, четвертой причиной появления широко иррадиированного возбуждения может быть механизм "обратной" афферентации, электрографическим выражением которого является как усиление реакции десинхронизации, так и появление вызванных потенциалов в первичных проекционных зонах "незаинтересованных" анализаторов в ответ на действие условных сигналов.
Как показали многочисленные опыты, по мере выработки и упрочения условной оборонительной реакции генерализованные изменения электрической активности начинают замещаться более локальными, очаговыми процессами. Однако таким локальным реакциям на всех стадиях формирования рефлекса (это будет проиллюстрировано в следующих главах) сопутствуют общие сдвиги, несколько более выраженные в первый момент действия сигнала условной реакции.
То, что генерализованные сдвиги ЭЭГ в стадии достаточно упроченных рефлексов не удается обнаружить при отведениях ЭЭГ от поверхности головы человека (Г. В. Гершуни, 1949; Е. Н. Соколов, 1955; Л. Г. Воронин, Е. Н. Соколов, 1955) или кролика (М. Н. Ливанов, 1960), объясняется особенностями записей накладными электродами, которые, как показали наблюдения Д. С. Воронцова (1960), И. С. Бериташвили (1960), А. И. Ройтбака (1958), А. Б. Когана (1960) и мн. др., богаты медленными колебаниями, в которых слабо отражаются диффузные быстрые процессы глубоких слоев коры (М. Я. Рабинович, 1961; Г. Джаспер и сотр., 1958; Ф. Моррелл, 1958б; X. Т. Чанг, 1958, и др.).
Диффузные процессы, сохраняющиеся, по нашим наблюдениям, даже в стадии вполне упроченных условных рефлексов, играют вполне определенную роль. В чем она состоит?
Генерализованные изменения электрической активности типа депрессии альфа-ритма Г. В. Гершуни (1949), Е. Н. Соколов (1955), Л. Г. Воронин и Е. Н. Соколов (1955) связывают с выражением ориентировочного рефлекса, который, по данным И. П. Павлова, а также П. К. Анохина (1949, 1958а), Е. Н. Соколова (1958, 1959), Л. Г. Воронина и Е. Н. Соколова (1955) и других, всегда сопутствует условному рефлексу, особенно на первых этапах его становления. В связи с этим общие условнорефлекторные сдвиги электрических потенциалов рассматривают как электрографическое выражение ориентировочной реакции.
Нам представляется, что генерализованная реакция ЭЭГ, сопутствующая условному рефлексу, вероятнее всего, является выражением совместного течения ориентировочного и условного рефлексов. И, действительно, еще А. И. Шумилиной (1945, 1949а, б, в) было показано, что диффузные процессы мозга обеспечивают собственно условнорефлекторные механизмы. А. И. Шумилиной удалось обнаружить, что деафферентированная конечность не только реагирует на всевозможные раздражения (Л. А. Орбели, К. И. Кунстман, 1924), определяемые диффузной афферентацией мозга, но и участвует в условнооборонительных реакциях здоровой конечности, причем ответы деафферентированной конечности опережают локальную оборонительную реакцию. Эти наблюдения позволили сделать вывод, что локальная условная двигательная реакция обеспечивается двумя формами возбуждения: диффузной и локальной (А. И. Шумилина, 1945, 1949а, б, в; П. К. Анохин, 1949). Диффузное возбуждение, сопутствующее локальному процессу, определяет, по мнению А. И. Шумилиной и П. К. Анохина, позиционную составляющую рефлекса, которая органически участвует в формировании временной связи.
Диффузное возбуждение ответственно не только за обеспечение ориентировочного рефлекса и позиционной компоненты условной реакции, оно также формируется тонически - обстановочным действием среды и появляется в обстановке эксперимента еще до применения условных раздражений. Последнее, как известно, создает функциональную настройку, известный фон для осуществления фазных реакций (П. С. Купалов, 1955; П. К. Анохин, 1949; Э. А. Асратян, 1959; Е. Н. Соколов, 1958, и др.).
Совершенно справедливо, что по диффузным процессам (синхронным изменениям ЭЭГ) нельзя судить о наличии временной связи (М. Н. Ливанов, 1960). Но сходство процессов в разных системах мозга при выработке условного рефлекса свидетельствует, как это показано М. Н. Ливановым и его сотрудниками, об установлении изохронизма в их деятельности. Изохронизм в деятельности систем, вступающих во временную функциональную связь, наблюдается в их корковых территориях (М. Н. Ливанов, 1958; В. Н. Думенко, 1955а, б; Т. А. Королькова, 1955), по поперечнику коры (И. Н. Книпст, 1955, 1958, 1960) и, как показали наши наблюдения, в специфических и неспецифических системах ствола мозга.
Таким образом, та констелляция нервных процессов, которая складывается на первых этапах формирования временной связи, включает широкий круг структур мозга.
Нами выделено две стадии генерализации, которые различаются как по показателям суммарной электрической активности, так и по показателям вызванных первичных ответов. Наличие двух периодов генерализации, выражающихся сначала в депрессии, а затем экзальтации электрических потенциалов, свидетельствует об изменении уровня условнорефлекторного возбуждения в процессе образования условного оборонительного рефлекса. Обе реакции, записанные биполярным способом с помощью погружных электродов, можно расценить как электрографическое выражение одного и того же процесса возбуждения. Очевидно, первый период связан с более высоким уровнем этого процесса, чем второй период. Об этом свидетельствуют наши наблюдения, показывающие, что при более интенсивной стимуляции наступает сдвиг разности потенциалов в сторону его депрессии и наоборот. О том же говорят и особенности динамики вызванных ответов, которые на первом этапе выработки рефлекса существенно уменьшаются. А, как известно, в период значительной активации мозга происходит уменьшение первичных потенциалов. Об этом же свидетельствует факт укорочения латентного периода вызванных первичных ответов в обеих стадиях.
Происхождение эффекта взаимодействия афферентаций на уровне каждой мозговой структуры, по-видимому, не является тождественным. Наличие условнорефлекторных изменений электрической активности в системе взаимодействующих анализаторов определяется встречей специфической и неспецифической афферентации, в то время как такие же изменения в "незаинтересованных" анализаторах, а также во всех других формациях мозга происходят вследствие взаимодействия лишь неспецифических афферентных влияний. Несмотря на то, что условнорефлекторные изменения активности в специфических и неспецифических системах мозга имеют разную природу, они находятся в коррелятивной связи, определяемой, очевидно, диффузной компонентой.
Естественно, встает вопрос, какое же значение имеет процесс взаимодействия возбуждений сигнального и безусловнорефлекторного происхождения на уровне каждого из образований мозга, ибо несомненно, коль скоро в этих образованиях имеются условнорефлекторные изменения ЭЭГ, следовательно, на их уровне осуществляется непосредственно или опосредованно через другие формации взаимодействие процессов, вызванных с анализаторов, вступающих во временную связь. И. П. Павлов установил, что формирование временной связи основано на встречном иррадиировании возбуждения из двух анализаторов, и полагал, что при упрочении рефлексов происходит проторение пути между их корковыми концами.
Исходя из этого положения школы И. П. Павлова и наблюдений, сделанных в последующие годы, следовало ожидать наиболее выраженных изменений активности в системе анализаторов, вступающих во временную связь. Однако такой вывод на основании изучения электрических процессов в стадии генерализации сделать не всегда можно. По-видимому, недостаточная информативность показателей электрической активности в этой стадии выработки рефлекса связана с маскировкой локальных процессов теми диффузными изменениями, которые широко охватывают мозг на этом этапе формирования реакции.
Вместе с тем, нередко удается видеть, что к концу периода генерализации на следах сочетаний обнаруживаются сдвиги активности, приуроченные к системам взаимодействующих анализаторов. Особенно отчетливо эти процессы выявляются в системе сигнального анализатора. Как уже отмечалось, две стадии генерализации сопровождаются сначала некоторым снижением, а затем возрастанием величины и длительности вызванных первичных ответов в слуховой корковой зоне (рис. 28). Эти наши наблюдения согласуются с ранее полученными данными В. В. Артемьева и Н. И. Безладновой (1952), В. В. Артемьева (1962), Galambos и сотр. (1956), Marsh и сотр. (1961), Р. С. Мнухиной (1961), Ван Тай-Ань и Н. И. Незлиной (1963) и др., показавшими нарастание потенциалов, вызываемых включением звуковых раздражений при превращении их в сигналы условной оборонительной реакции, на других экспериментальных животных (кошках, обезьянах).
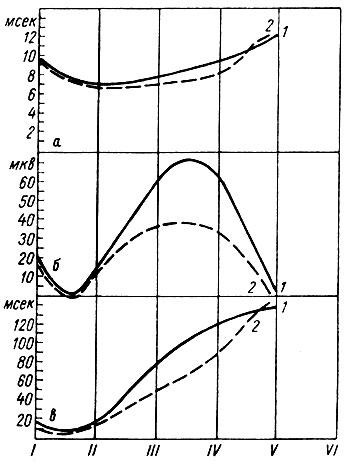
Рис. 28. Схема соотношения изменений латентного периода, амплитуды и длительности первичных вызванных потенциалов в корковой проекционной зоне сигнального (слухового) анализатора в процессе становления условного оборонительного рефлекса собак. а -динамика латентного периода; б - динамика амплитуды; в - динамика длительности первичного ответа. 1 - ответ на 1-й; 2 - ответ на 3-й щелчок. По оси абсцисс - стадии формирования рефлекса; по оси ординат на а, в - миллисекунды, на б - микровольты (по Т. С Наумовой и Н. С. Поповой, 1967б)
Как и в опытах В. В. Артемьева и Н. И. Безладновой (1952), повышение амплитуды вызванных ответов на первые щелчки в серии происходит не сразу. Выработка рефлекса начинается с периода, когда вызванные потенциалы имели тот же характер, что и до начала сочетаний. Этот период совпал с первой фазой формирования условного оборонительного рефлекса - фазой генерализованной депрессии электрических потенциалов.
Следует указать на то, что отмеченная закономерность обнаружилась в поведении амплитудной характеристики как позитивного и негативного компонента первичного ответа, так и позитивного компонента вторичного ответа, а не только поздних компонент ответа (И. А. Пеймер, 1958, 1959; Worden, 1959, и др.).
Как показали наши данные, латентный период таких ответов в I и во II стадиях образования рефлекса укорачивается по сравнению с величиной этого параметра до выработки рефлекса (см. рис. 28). Это значит, что условный сигнал встречает состояние известной подготовленности анализатора к его действию, т. е. состояние повышенной возбудимости. Последнее было обнаружено А. А. Князевой и И. Э. Барбель (1952), А. А. Гюрджан (1953), А. М. Марусевой и Л. А. Чистович (1954), Е. Н. Соколовым (1960) и др. Так, А. А. Князева и И. Э. Барбель (1962) нашли изменение чувствительности зрительного анализатора под влиянием сочетания света с болевым подкреплением у человека. А. А. Гюрджан (1953) обратил внимание на расширение границ поля зрения для световых раздражений, становящихся сигналом условной реакции. В исследованиях А. М. Марусевой и Л. А. Чистович (1954), Hughes (1958) было показано на людях понижение порога условной реакции и на звуковые раздражения. Повышение чувствительности сигнального анализатора происходит и в отношении ориентировочных реакций (Е. Н. Соколов, 1959). Е. Н. Соколов связывает изменения возбудимости сигнального анализатора главным образом с ориентировочной реакцией, в то время как П. О. Макаров (1960) считает, что в сигнальном анализаторе меняется возбудимость, лабильность и структура условного рефлекса. О механизме этого влияния речь будет идти в следующей главе.
Как подчеркивалось, величина вызванных потенциалов в первичной проекционной корковой зоне слуха в большой мере зависит от порядкового номера щелчка в серии, на которую вырабатывается ответная реакция. Это выражается в том, что вызванные ответы по мере приближения к моменту условнорефлекторной реакции значительно редуцируются. Падение амплитуды первичного ответа на ритмические щелчки по мере приближения к моменту условной реакции отмечалось всеми исследователями, анализировавшими условнорефлекторные изменения вызванных потенциалов (А. И. Ройтбак, 1958; А. Б. Коган, 1960; Г. Джаспер и сотр., 1958; Г. Д. Смирнов, 1960; Л. Г. Трофимов и Н. Н. Любимов, 1960; Hearst и сотр., 1960; Marsh и сотр., 1961, и др.). Bremer (1938), а затем В. В. Артемьевым (1951), А. И. Ройтбаком (1960) и другими было замечено, что при возбужденном состоянии животного первичные ответы обычно не наблюдаются, поэтому Bremer, Bonnet (1950), Г. Д. Смирнов (1960) считают, что исчезновение вызванных ответов связано с маскировкой этой реакции. А. И. Ройтбак (1960) относит их отсутствие за счет тормозного нисходящего влияния ретикулярной формации на проведение возбуждения по специфическим путям анализатора условного сигнала. А по данным Г. Джаспера и сотр. (1958), уменьшение вызванных ответов в специфической сенсорной области до и во время условной реакции объясняется переходом возбуждения от сенсорного звена условного рефлекса к моторному. Такое объяснение было выдвинуто М. Н. Ливановым и К. Л. Поляковым (1945), М. Н. Ливановым и сотрудниками (1951), но для факта исчезновения вызванных ответов из коркового представительства сигнального анализатора в стадии концентрации рефлекса. А. Б. Коган (1960) полагает, что исчезновение вызванных ответов при осуществлении рефлекса происходит за счет перемещения очагов активности в слоях коры, так как полярность ответов (при биполярном способе отведения) в слоях меняется. И, наконец, Е. Н. Соколов (1959), Jus и Jus (1959а, б) относят изменение специфически вызванных потенциалов и общей реакции "arousal", к состоянию ориентировочного рефлекса.
Анализ наших данных показал, что уменьшение вызванных потенциалов к моменту условной оборонительной реакции происходит при одновременном укорочении их латентного периода и длительности (см. рис. 28) (ответ на 3-й щелчок). Укорочение латентного периода и длительности первичных ответов объясняется повышением скорости возникновения и проведения условнорефлекторного возбуждения, которые меняются в течение действия условного сигнала. Мы полагаем, что облегчение возникновения и проведения этого возбуждения может быть объяснено повышением возбудимости и лабильности сигнального анализатора в процессе подготовки и осуществления условной реакции, а также, возможно, включением в проведение условнорефлекторного возбуждения путей с меньшим количеством переключений по их ходу.
Результаты измерения латентных периодов и длительности ответов позволяют присоединиться к взгляду, что исчезновение вызванных потенциалов в момент осуществления условного рефлекса объясняется эффектом маскировки, связанным с ускоренным возникновением и протеканием вызванных потенциалов (Г. Д. Смирнов, 1960). Менее вероятно, что редукция вызванных ответов при осуществлении эффекторной реакции связана с переходом возбуждения от сенсорного звена условного рефлекса к моторному, как это полагают Г. Джаспер и сотр. (1958), так как в этот период происходит исчезновение как первичных, так и вторичных вызванных ответов из всех зон мозга (в том числе сенсомоторных).
Взаимодействие сигнального и безусловнорефлекторного возбуждений в стадии генерализации осуществляется в четкой форме и на уровне коркового конца кожно-двигательного анализатора, где оно имеет превалирующий характер. Об этом свидетельствует эффект локальной активации сенсомоторной зоны коры в участке представительства реагирующей конечности, его уровень превышает уровень прироста активности в других структурах мозга. Об этом же говорит значительное увеличение амплитуды вызванных ответов в сенсомоторной зоне коры в обеих стадиях генерализации по сравнению с фоновыми реакциями, с одной стороны, и по сравнению с ответами в сигнальном анализаторе, - с другой.
Еще более выраженные изменения электрических потенциалов в системе взаимодействующих анализаторов обнаруживались при дальнейшей выработке рефлекса. Мало вероятно, чтобы активная роль анализаторов приобреталась ими только в стадии концентрации, поэтому есть все основания считать, что и в стадии генерализации рефлекса определяющее значение в замыкании имеет взаимодействие процессов, осуществляемое в самих анализаторах, вступающих во временную связь. На этом основании можно думать, что в стадии генерализации условного рефлекса поведение основных компонент реакции не определяется взаимодействием процессов на уровне ретикулярной формации, ибо условнорефлекторные изменения ЭЭГ характерны не только для этой формации, но и для других зон мозга и в первую очередь анализаторов, вступающих во временную связь. Так как эти изменения имеют место как на уровне их корковых концов (Durup и Fessard, 1935; М. Н. Ливанов и К. Л. Поляков, 1945; М. Н. Ливанов и А. М. Рябиновская, 1947; М. Н. Ливанов и Т. А. Королькова, 1948; В. В. Артемьев, Н. И. Безладнова, 1952; Р. И. Лурье и сотр., 1956, и др.), так и на уровне их первичных переключательных реле (М. Н. Ливанов и сотр., 1951; Galambos и сотр., 1956; Hernandez-Peon и сотр., 1956а; наши наблюдения и др.), трудно думать, что они определяются ретикулярными влияниями. Вместе с тем, рефлекс представляет собой целостную реакцию организма, включающую и соматические, и вегетативные, и эндокринногуморальные компоненты (И. П. Павлов; П. К. Анохин, 1949, 1958а, и др.). Естественно, что все эти реакции идут с участием различных мозговых структур. Только тем, что в ориентировочных и условных реакциях организма участвуют различные структуры мозга, можно объяснить весь тот сложный комплекс процессов, которым определяется реакция организма как целого. Очевидно, целостная реакция организма и определяется взаимодействием сигнальной и подкрепляющей афферентации на уровне самых различных систем мозга, в том числе на уровне ретикулярной формации ствола и других неспецифических структур мозга, а также специфических систем других модальностей. Последнее, как уже отмечалось, играет немаловажную роль в повышении уровня возбуждения взаимодействующих анализаторов.
Уже указывалось, что констелляция процессов, включающая в стадии генерализации большинство структур мозга, возникает сначала на следах очередных сочетаний, а затем воспроизводится условнорефлекторно. Нельзя не подчеркнуть, что формирующаяся констелляция имеет доминантные свойства и воспроизводится при всякого рода посторонних раздражениях, равно как и сопровождает межсигнальные реакции. О природе последних более подробно речь идет в главе V.
Изучение следовых реакций в хронических опытах проводилось лишь в отношении незначительного числа образований, среди которых отсутствуют подкорковые отделы специфических систем анализаторов, вступающих во временную связь.
Как показали наши наблюдения, сохранение следов возбуждения при выработке оборонительных условных рефлексов на звук не является привилегией лишь кортикальных, ретикулярных и других поливалентных систем мозга. На первых этапах выработки оборонительного условного рефлекса на звуковые раздражения изменения ЭГ стволовых и корковых структур в последействии условного раздражения имели генерализованный характер. Эти данные позволяют утверждать, что следовые процессы на первых этапах формирования условной реакции охватывают все исследованные структуры мозга.
Итак, приведенные выше наблюдения не оставляют сомнения в том, что условнорефлекторные изменения электрических процессов и сохранение их следов имеют место не только в коре ретикулярной формации и других неспецифических ядрах, но также на уровне всех систем мозга.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'