I стадия формирования условного оборонительного рефлекса (первый период генерализации)
По нашим наблюдениям, диффузные изменения основной электрической активности носят различный характер в разные периоды стадии генерализации. В первый период генерализации действие условного сигнала вызывает снижение амплитуд и ритма электрических колебаний (депрессии активности) в стволово-подкорковых образованиях и корковых формациях мозга, ЭГ которых отводится с помощью погружных электродов. Если запись электрической активности коры проводится накладными электродами, в ней наблюдается или снижение величины и возрастание частоты электрических осцилляций или увеличение этих же показателей.
Как уже указывалось, электрокожное раздражение до выработки рефлекса сопровождалось либо генерализованным по подкорково-стволовым и норковым структурам снижением, либо таким же возрастанием электрической активности, так как, по мере привыкания животного к болевому воздействию, характер электрографической реакции менялся, до начала выработки рефлекса обычно устанавливалась более или менее стационарная картина, которая, как правило, представляла реакцию экзальтации активности, генерализованной по подкорково-стволовым и кортикальным структурам. В начале выработки рефлекса описываемая реакция сохранялась только на действие безусловного стимула.
У большинства собак после нескольких сочетаний звукового и болевого раздражений условный сигнал начинал вызывать не генерализованную экзальтацию активности, которая возникала при пороговой болевой стимуляции до выработки рефлекса, а ту, которая регистрировалась при действии достаточно сильных электрокожных стимулов, - реакцию снижения активности. Именно эти условнорефлекторные изменения электрических потенциалов регистрируются сначала на следах первых сочетаний, а затем в ответ на действие условного сигнала, опережая появление оборонительного рефлекса.
Обращает на себя внимание в этот период выработки рефлекса значительное сходство электрических реакций в так называемых специфических и неспецифических системах каждого уровня нервной системы.
Угнетение электрических колебаний наступало прежде всего в структурах продолговатого мозга (его латентный период зависел от индивидуальных особенностей нервной системы животного, а также количества предшествующих сочетаний), а затем на уровне среднего мозга и в субкортикальных и кортикальных образованиях. Скорость распространения процесса депрессии зависела от количества предшествующих сочетаний и наличия или отсутствия эффекторной реакции. Чем ближе был момент появления условного рефлекса, тем выше была скорость распространения этого процесса в направлении от продолговатого мозга к коре, т. е. тем меньшие различия удалось наблюдать в латентном периоде наступления электрографического ответа стволовых, подкорковых и корковых структур. На рис. 13 представлены записи ЭГ разных зон коры левого полушария и структур продолговатого мозга справа (кохлеарных ядер, крупноклеточного ядра ретикулярной формации продолговатого мозга) собаки № 18 в начальный период выработки условного оборонительного рефлекса (7-е сочетание подпорогового тона 500 гц 40 дб и электрокожного раздражения правой передней лапы). Звуковой сигнал при его 7-м применении (рис. 13), когда условная двигательная реакция еще отсутствовала, сначала не вызывал никаких изменений электрической активности. Через 980 мсек его изолированного действия наблюдалось незначительное снижение амплитуд электрических колебаний кохлеарных ядер, крупноклеточного ядра ретикулярной формации, а еще через 580 мсек снижение активности наступало в корковых концах слухового, а затем и кожно-двигательного анализаторов. В последнем падение амплитуд колебаний происходит почти вдвое (с 30 до 12-15 мкв), в то время как их доминирующий ритм возрастал в то же число раз. Снижение амплитуд колебаний не сопровождается заметными изменениями частотного показателя активности ни в кохлеарных ядрах, ни в слуховой зоне коры большого мозга. В остальных отведениях (от коры латеральной и супрасильвиевой извилин большого мозга) каких-либо существенных изменений ЭЭГ отметить не удается. Действие порогового электрокожного раздражения вызывает генерализованное нарастание амплитуд и частот электрических колебаний, как и до выработки рефлекса. Продолжительность I стадии выработки рефлекса у этого животного - около 40 сочетаний.
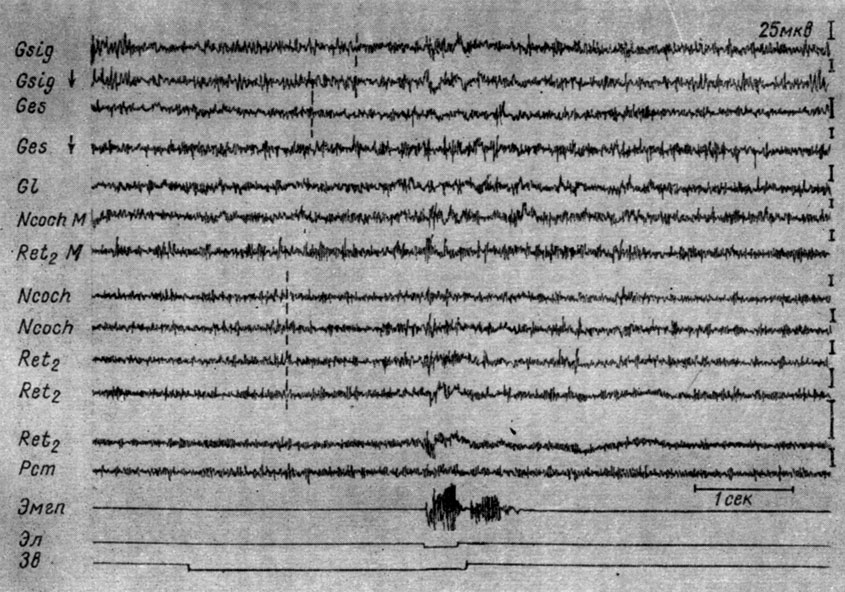
Рис. 13. Изменения ЭГ разных образований коры большого мозга и продолговатого мозга собаки № 18, наступающие в I стадии формирования условного оборонительного рефлекса. 7-е сочетание тона 500 гц 40 дб с электрокожным раздражением правой передней лапы (опыт № 7 от 13/VIII 1960 г.). Начало эффекта депрессии электрической активности в структурах мозга отмечено пунктиром
Пример изменений ЭГ на уровне продолговатого мозга и разных зон коры в I стадии представлен на рис. 14. На этом рисунке приводятся ЭГ разных зон коры левого полушария и сенсомоторной зоны коры правого, а также структур продолговатого мозга во время 91-го применения порогового тона 500 гц 48 дб, сочетающегося с электрокожным пороговым раздражением правой лапы у собаки № 4. Отчетливо видно, что звук вызывает в структурах продолговатого мозга урежение колебаний электрической активности (с 55-60 в 1 сек до 40-45 в 1 сек), которому через 300-400 мсек после включения звукового сигнала начинает сопутствовать снижение амплитуд колебаний до нескольких микровольт. Обращает на себя внимание близость изменений электрической активности в ядрах нежного и клиновидного канатика, кохлеарных ядрах и нижнем центральном ядре ретикулярной формации. Снижение электрической активности в этих ядрах наступает одновременно, но восстановление исходной частоты и амплитуд колебаний происходит в кохлеарных ядрах раньше, чем в ядрах нежного и клиновидного канатиков. Совершенно так же, как в кохлеарных ядрах, меняются при записи погружными электродами электрические процессы в сенсомоторной зоне коры. Однако снижение уровня активности здесь происходит на 150-200 мсек позднее, чем в структурах ствола, и ему предшествует вспышка активности. Ничего похожего не удается наблюдать в записи электрической активности коры сигмовидной, сильвиевой, супрасильвиевой и латеральной извилин, сделанных накладными электродами. Во всех этих отведениях происходит возрастание ритма колебаний (с 40-45 до 50-55 в 1 сек) при неизменной (в эктосильвиевой, супрасильвиевой извилинах) или возрастающей (в правой и левой сигмовидной и латеральной извилинах) амплитуде колебаний. Обращает внимание резкое (более чем в 2 раза) возрастание уровня электрической активности левой сенсомоторной зоны коры, совпадающее с периодом наиболее глубокой депрессии активности в стволовых структурах и движением конечности.
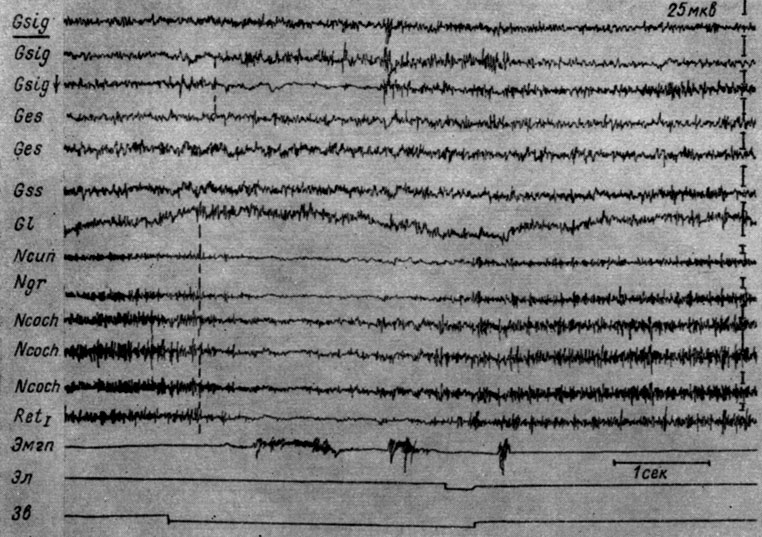
Рис. 14. Изменения ЭГ разных образований коры большого мозга и продолговатого мозга собаки № 4, наступающие в I стадии образования условного оборонительного рефлекса. 91-е сочетание тона 500 гц 48 дб с электрокожным раздражением правой передней лапы (опыт № 19 от 30/I 1959 г.). Пунктиром отмечено начало реакции депрессии
На рис. 15, а видно, что период генерализованной депрессии электрических потенциалов у этого животного длился более 100 сочетаний. Некоторые сочетания сопровождаются изменениями ЭГ, характерными для следующих стадий формирования рефлекса. В момент, предшествующий и сопутствующий двигательной реакции, в коре сенсомоторной зоны, записываемой накладными электродами, регистрируется эффект экзальтации электрических колебаний (см. рис. 15, б), однако прирост активности в этой зоне мозга на этом этапе формирования условной реакции несколько меньше, чем на последующих этапах.
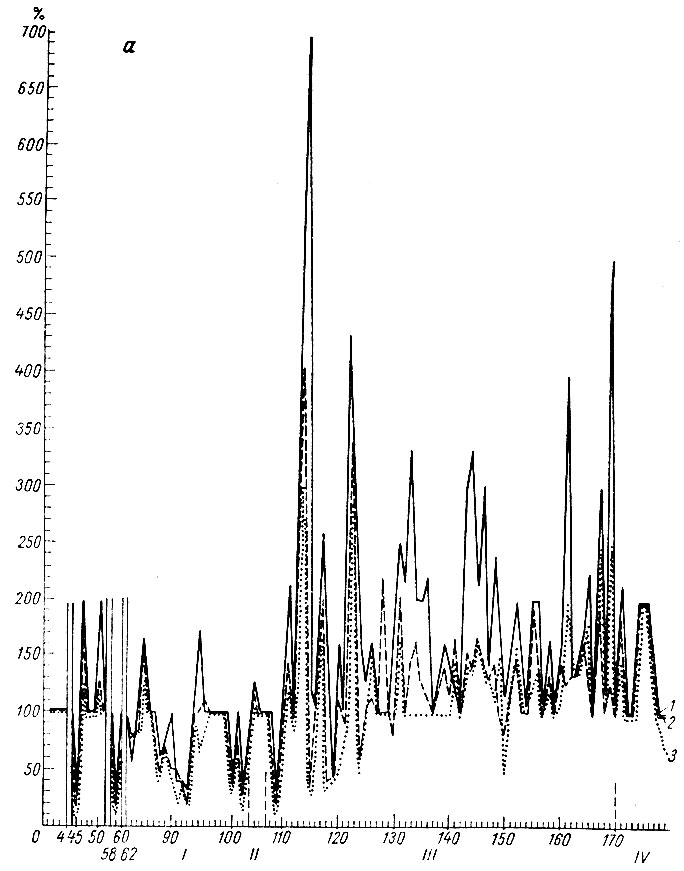
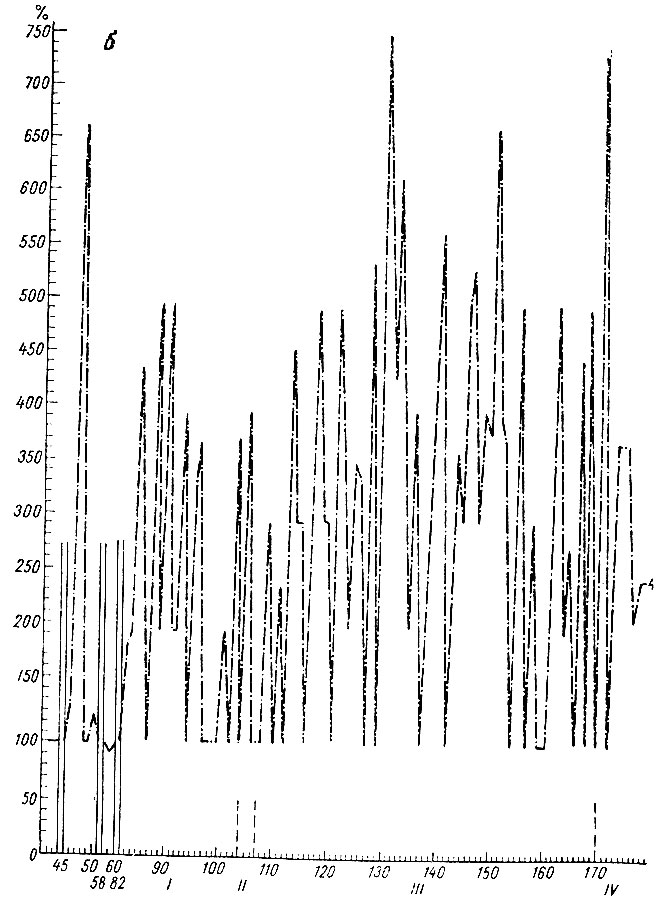
Рис. 15. Амплитудная характеристика ЭГ условной реакции при выработке оборонительного рефлекса на тон 500 гц 48 дб (собака № 4). a: 1 - ЭГ кохлеарного ядра; 2 - ЭГ ядра нежного и клиновидного канатиков; 3 - ЭГ ретикулярной формации продолговатого мозга (нижнее центральное ядро); б: 4 - ЭГ сигмовидной извилины. По оси абсцисс - порядковые номера сочетаний и стадий образования рефлекса; по оси ординат - проценты по отношению к исходным величинам
Сопоставление изменений суммарной электрической активности с изменениями амплитуды первичных ответов в слуховой зоне той же собаки № 4 (рис. 16) на включение условного сигнала - тона 500 гц 48 дб - показало, что в первой фазе формирования условной оборонительной реакции (1-106 сочетания) происходит существенное снижение величины вызванных потенциалов.
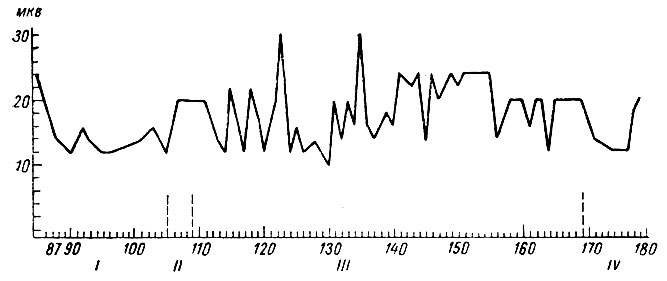
Рис. 16. Динамика амплитуды первичных вызванных потенциалов на включение условного сигнала оборонительного рефлекса - тона 500 гц 48 дб (собака № 4). По оси абсцисс - номера сочетаний и стадий формирования рефлекса; по оси ординат - микровольты
Как видно на рис. 17 (а, б), следовые изменения электрической активности и в стволовых и кортикальных структурах на I стадии формирования условного оборонительного рефлекса у собаки № 4 сопровождают почти каждое сочетание. В начале этой стадии следовые процессы выражаются в генерализованной депрессии электрических колебаний, а после 80 сочетаний либо в генерализованной экзальтации их, либо в избирательной активации процессов в кохлеарных ядрах и сенсомоторной зоне (кора сигмовидной извилины). Следовые изменения ЭГ, наблюдающиеся в конце I стадии, отражают сдвиги, которые сопутствуют условной реакции на следующих этапах образования рефлекса. Длительность следовых процессов в этой стадии выработки рефлекса не превышает у этого животного 25 сек (рис. 18).
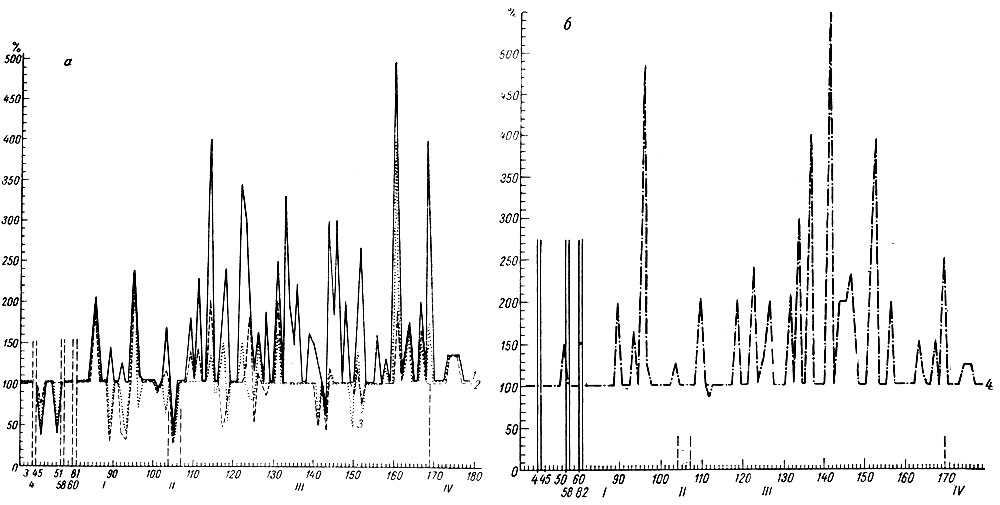
Рис. 17. Амплитудная характеристика ЭГ последействия условного оборонительного рефлекса на тон 500 гц 48 дб (собака № 4). а - обозначения те же, что на рис. 15, а; б - обозначения те же, что на рис. 15, б
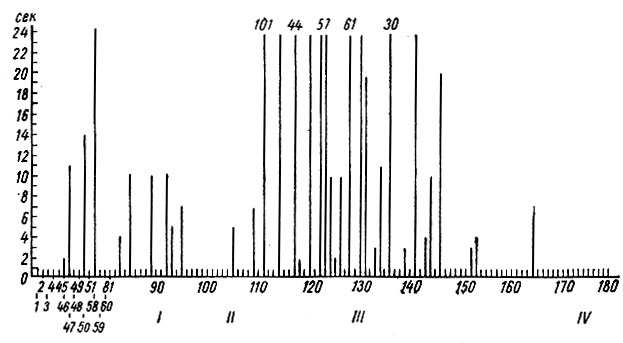
Рис. 18. Длительность ЭГ последействия условного оборонительного рефлекса на тон 500 гц 48 дб (собака № 4). По оси абсцисс - порядковые номера сочетаний и стадий выработки рефлекса; по оси ординат - секунды
Измерения величины вызванных потенциалов (отводимых от коры с помощью накладных электродов) на включение условного сигнала (тона 500 гц 48 дб) показали, что на протяжении первой стадии ответы по-разному меняются в первичной проекционной слуховой и сенсомоторной зонах коры головного мозга, а именно, они слабо изменяются (уменьшаются) в коре эктосильвиевой извилины и резко возрастают в коре сигмовидной извилины головного мозга у собаки № 3 (рис. 19).
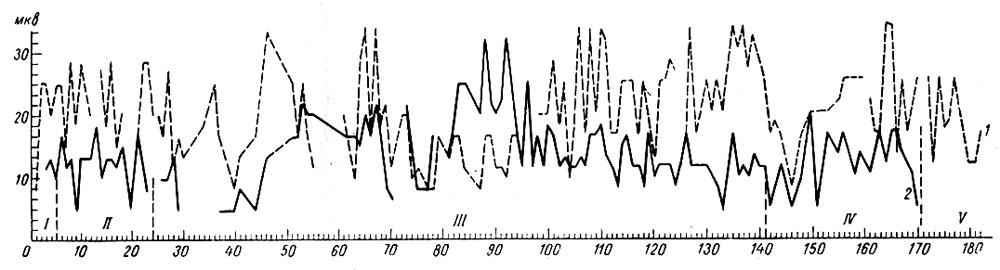
Рис. 19. Динамика амплитуды вызванных потенциалов в коре сигмовидной (1) и эктосильвиевой (2) извилин на включение условного сигнала оборонительного рефлекса - тона 500 гц 48 дб (собака № 5). По оси абсцисс - порядковые номера сочетаний и стадий формирования рефлекса, по оси ординат - микровольты
Электрические процессы, регистрируемые в разных структурах мозга с помощью погружных электродов, при осуществлении условных оборонительных рефлексов на прерывистые и непрерывные звуковые стимулы, по интенсивности значительно превышающие пороговые значения, имели много общего с теми процессами, которые регистрировались при действии сигналов подпороговой и пороговой интенсивности, а именно вызывали генерализованное снижение электрической активности. В случае ритмических сигналов (щелчков) на фоне развивающейся депрессии колебаний потенциалов могут регистрироваться и вызванные ответы в разных структурах мозга. Так, у собаки № 4 (рис. 20) выработка условного оборонительного рефлекса осуществлялась и на прерывистые (7 в 1 сек) щелчки интенсивностью 56 дб. Формирование рефлекса происходило после упрочения условной реакции на сигнал 500 гц 48 дб. Включение ритмической стимуляции сопровождалось сначала незначительным снижением общего уровня активности, на фоне которого в кохлеарных ядрах, ретикулярной формации появились вызванные потенциалы. В период максимальной их выраженности возник мощный эффект экзальтации электрических колебаний в левой сенсомоторной зоне коры мозга, на фоне которого в кохлеарных ядрах развилось, а в остальных ядрах продолговатого мозга углубилось снижение электрической активности, предшествующее и совпадающее с появлением токов действия на ЭМГ. Электрокожное раздражение существенно не изменило эту картину активности. Следовая экзальтация активности длилась более 2 сек.
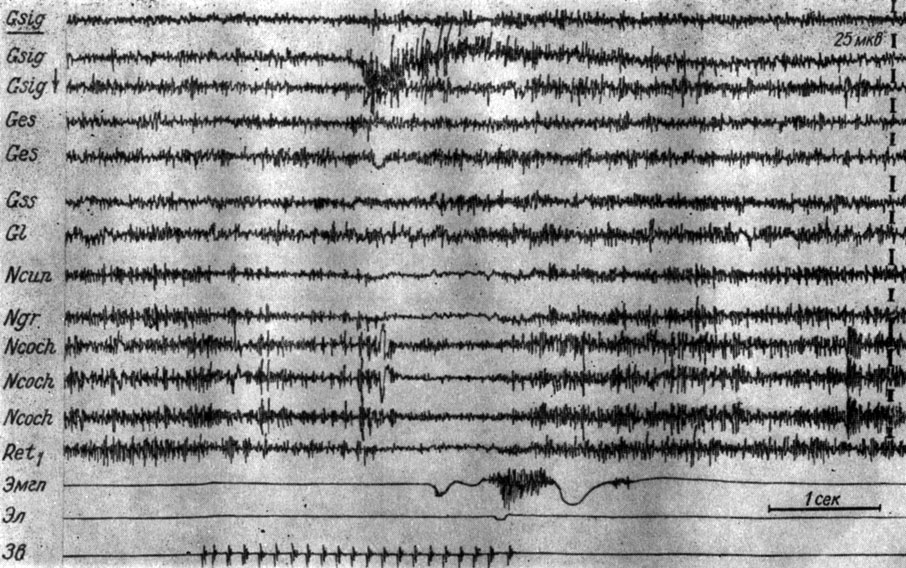
Рис. 20. Изменения ЭГ коры большого мозга и образований продолговатого мозга собаки № 4, наступающие в I стадии формирования условного оборонительного рефлекса на звуковые щелчки (7 в 1 сек, 56 дб). 31-е сочетание щелчков с электрокожным раздражением правой передней лапы (опыт № 54 от 3/IV 1959 г.)
Анализ вызванных ответов на звук проводился в отношении потенциалов, зарегистрированных на уровне первичной корковой слуховой проекционной зоны.
Анализ параметров (латентного периода, амплитуды и длительности) первых трех компонент первичных вызванных потенциалов на условные ритмические (2 в 1 сек) звуковые раздражения (щелчки) показал, что в I стадии формирования рефлекса происходят известные изменения. Сказанное иллюстрируют рис. 21-24 (собака № 19). На рис. 21, а представлены ответы в первичной проекционной зоне слуха, наступающие при действии звуковых щелчков до выработки рефлекса, и на рис. 21, б в первый период формирования условной оборонительной реакции на них. Хотя каких-либо различий в величине вызванных ответов до и в начале образования условного рефлекса заметить не удается, однако тщательный анализ величины каждого компонента ответа выявил в ряде случаев снижение их величины. На рис. 22 по оси абсцисс отложены порядковые номера сочетаний, по оси ординат - величины амплитуд вызванных ответов на 1-й (1), 3-й (2) и 4-й (3) щелчки (из серии в 6 щелчков, служащей сигналом условной реакции) в слуховой области коры больших полушарий головного мозга той же собаки. Сверху вниз расположены графики динамики изменения амплитудного максимума первой положительной, максимума отрицательной и, наконец, максимума второй позитивной компоненты ответа. Из этого графика следует, что условный ритмический стимул либо не вызывал существенных изменений величины компонент ответа, либо сопровождался значительным снижением амплитуды вплоть до полного их исчезновения (например, первой и второй позитивной компоненты ответа).
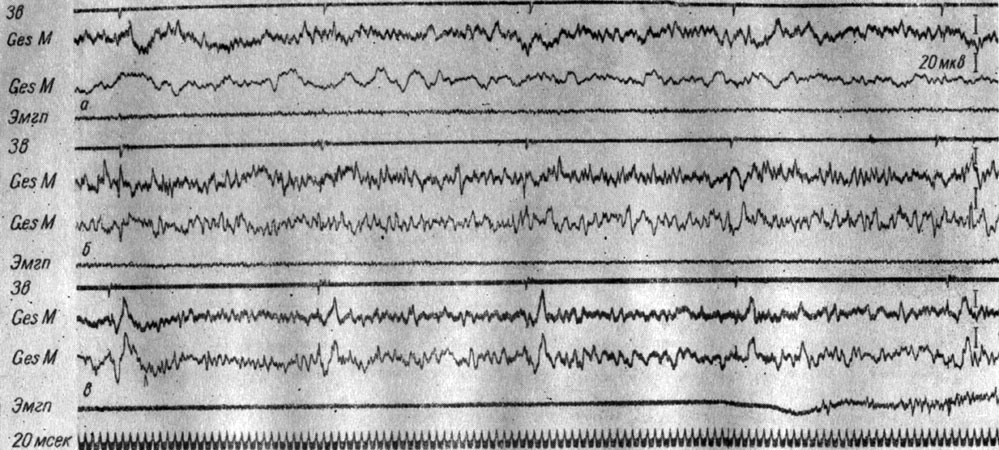
Рис. 21. Изменения вызванных потенциалов в разных участках первичной проекционной корковой зоны слуха собаки № 19 в I (б) и II (в) стадиях образования условного оборонительного рефлекса на ритмические (2 в 1 сек) звуковые раздражения (щелчки 56 дб). а - до выработки условного рефлекса (опыт № 1 от 24/IV 1964 г.); б - 4-е сочетание (опыт № 3 от 6/V 1964 г.); в - 25-е сочетание (опыт № 12 от 20/V 1964 г.) (по Т. С. Наумовой, Н. С. Поповой и Н. Н. Ульяниной, 1966 г.)
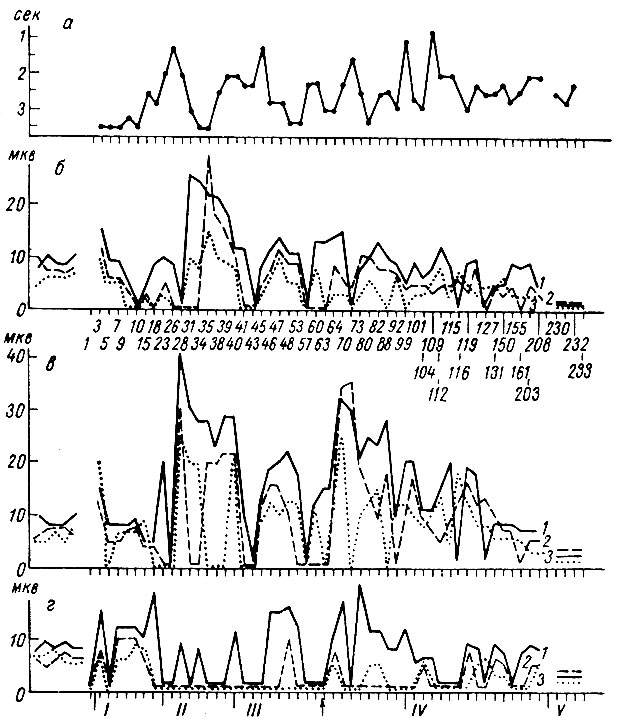
Рис. 22. Динамика латентного периода эффекторной реакции (а) и амплитуды разных фаз первичных вызванных потенциалов (б, в, г) при образовании оборонительного условного рефлекса на щелчки 2 в 1 сек, 56 дб (собака № 19). б - амплитуда первой позитивной; в - негативной и г - второй позитивной фазы первичного ответа. 1 - амплитуда ответа на 1-й щелчок; 2 - на 3-й щелчок; 3 - на 4-й щелчок. По оси абсцисс - порядковые номера сочетаний и стадий выработки оборонительного рефлекса; по оси ординат - на а - секунды, на б, в, г - микровольты. Стрелка - момент введения дифференцировки (по Т. С. Наумовой, Н. С. Поповой и Н. Н. Ульяниной, 1966)
Латентный период (рис. 23) и длительность (рис. 24) тех же ответов у собаки № 19 также претерпевают выраженное снижение. Описанные сдвиги параметров трех компонент вызванного ответа касаются реакций на первый, третий и четвертый щелчки.
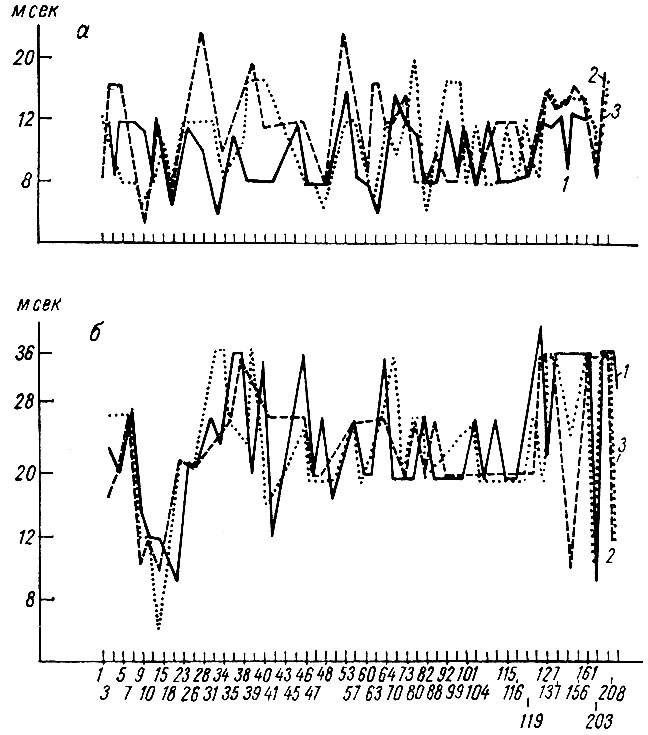
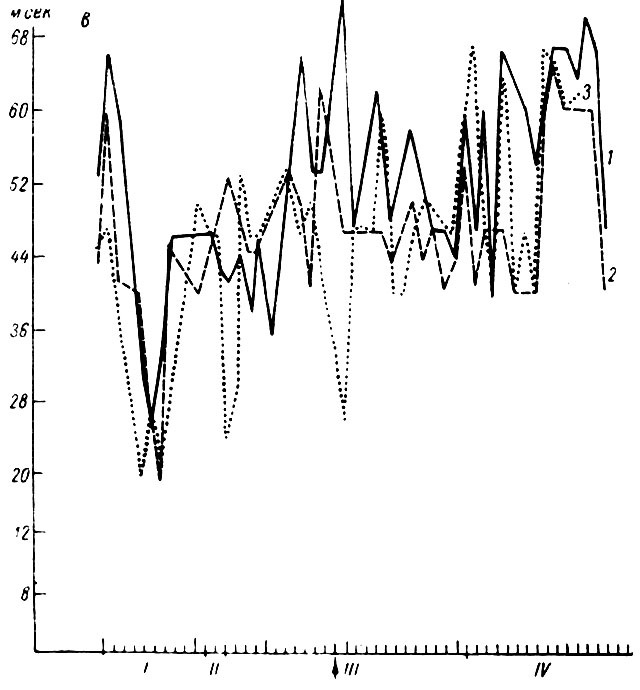
Рис. 23. Динамика латентного периода первичных вызванных потенциалов при выработке оборонительного условного рефлекса на щелчки 2 в 1 сек (собака № 19). а - латентный период первичного ответа; б - время до максимума первой позитивной; в - время до максимума негативной фазы ответа, 1 - латентный период и время до максимума позитивной и негативной фазы на 1-й щелчок; 2 - на 3-й щелчок; 3 - на 4-й щелчок. По оси абсцисс - порядковые номера сочетаний и стадий образований рефлекса; (на этом и последующих аналогичных рисунках номера сочетаний относятся и к в, а номера стадий и к а, б); по оси ординат - миллисекунды. Стрелка - момент введения дифференцировки (по Т. С. Наумовой и Н. С. Поповой, 1967а)
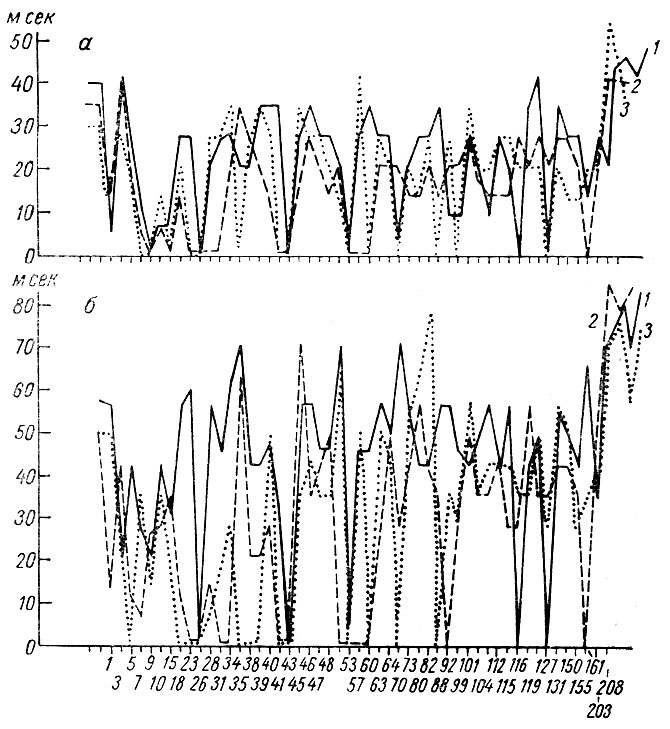
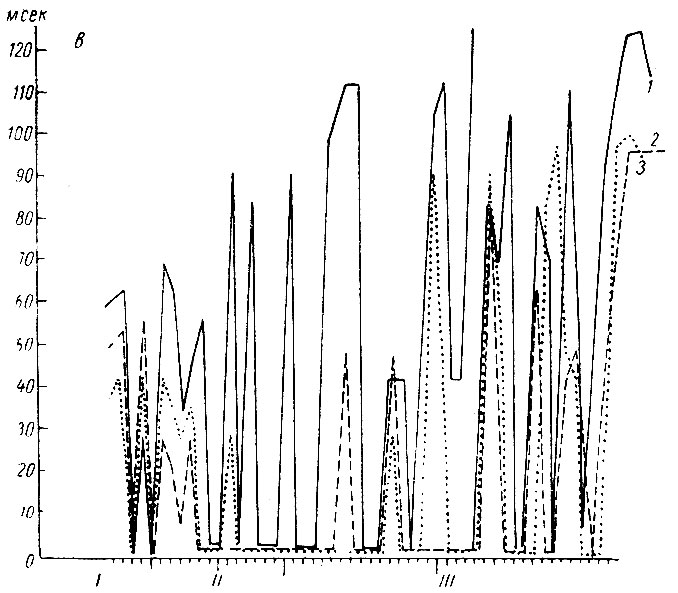
Рис. 24. Динамика длительности разных фаз первичных вызванных потенциалов при выработке условного оборонительного рефлекса на щелчки 2 в 1 сек 56 дб (собака № 19). а - длительность первой позитивной; б - негативной; в - второй позитивной фазы первичного ответа. 1 - длительность компонент ответа на 1-й щелчок; 2 - на 3-й щелчок; 3 - на 4-й щелчок. По оси абсцисс и ординат - то же, что на рис. 23. Стрелка - момент введения дифференцировки (по Т. С. Наумовой и Н. С. Поповой, 1967 б)
Итак, условная реакция на звуковой сигнал на I этапе выработки рефлекса формируется на фоне генерализованной депрессии (снижения частоты и амплитуд колебаний) электрической активности, обнаруженной в записях погружными электродами на разных уровнях мозга: кохлеарных ядрах, ядрах нежного и клиновидного канатиков, внутреннем коленчатом теле, заднем и переднем двухолмии, ядрах ретикулярной формации ствола мозга, красном ядре, черной субстанции, центральном сером веществе, вентральном и ретикулярном ядрах: таламуса, субталамическом ядре, гипоталамусе, структурах обонятельного мозга (гиппокампе, миндалевидном ядре), хвостатом ядре и, наконец, территориях новой коры. Эта форма активности появляется сначала только на следах сочетаний, а затем и в ответ на действие условного звукового сигнала. Те же изменения активности регистрируются и в межсигнальные периоды "спонтанно" или в ответ на посторонние внешние раздражения. На следах сочетаний к концу I стадии появляются изменения электрической активности, характерные для II и III стадий формирования рефлекса.
Обнаруживается четкая корреляция глубины реакции депрессии и латентного периода ее появления в образованиях каждого уровня мозга: продолговатого мозга (кохлеарных ядрах, верхнем оливарном комплексе ядер, трапециевидном теле, ядрах нежного и клиновидного канатика, нижнем центральном, мелкоклеточном и крупноклеточном ядрах ретикулярной формации и др.), среднего мозга (ядрах латеральной петли, внутреннем коленчатом теле, заднем двухолмии, ядрах ретикулярной формации среднего мозга - nucl. cuneiformis, nucl. subcuneiformis, красном ядре, черной субстанции, центральном сером веществе и др.), межуточного мозга (центральном и ретикулярном ядрах таламуса, субталамическом ядре, ядрах гипоталамуса и др.) и т. д., т. е. анализаторных и, условно говоря, межанализаторных или специфических и неспецифических формациях ствола мозга.
Анализ параметров (латентного периода, амплитуды и длительности) первых трех компонент вызванных потенциалов, регистрируемых в первичной проекционной зоне слуха накладными электродами, на включение условных звуковых раздражений или серию щелчков показал, что в I стадии формирования оборонительного условного рефлекса эти параметры снижаются по сравнению с теми же показателями ответов на действие тех же стимулов до выработки рефлекса.
Стадия генерализованной депрессии активности у разных животных имела разную продолжительность: у одних собак она была мимолетной, и ее трудно было уловить (таких у нас было 6 собак), а у других (17 животных) она колебалась от 5 до 100 сочетаний, что зависело от индивидуальных особенностей нервной системы подопытных животных (табл. V).
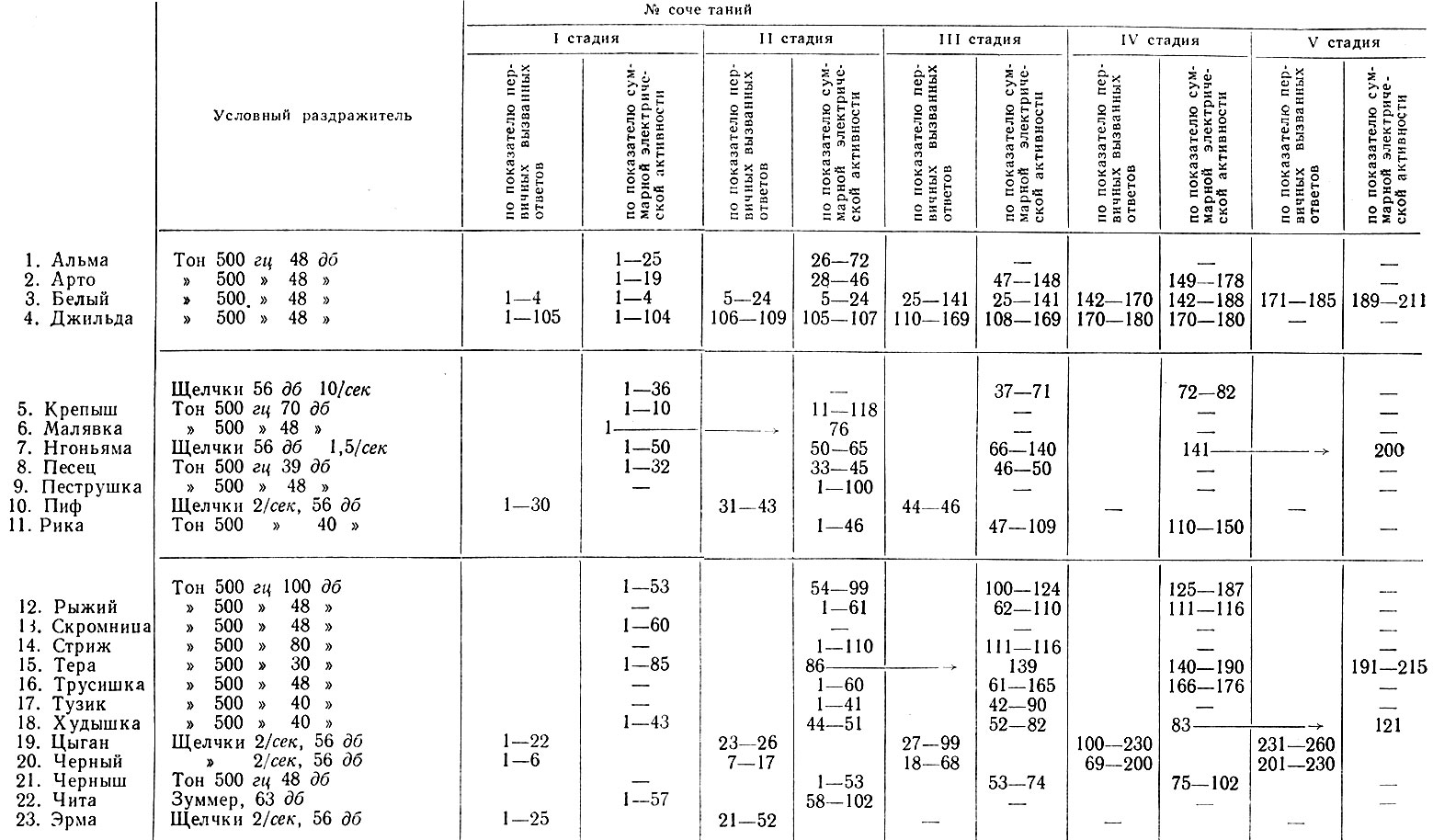
Таблица V. Продолжительность стадий формирования условного оборонительного рефлекса у подопытных собак
Примечание. Стрелки указывают, что нет данных для отнесения указанной серии сочетаний к одной из тех стадий, между которыми стоит стрелка.
На последующих стадиях выработки рефлекса, вплоть до периода их значительного упрочения, мог наблюдаться возврат к первому этапу генерализации, причем у части собак это явление наблюдалось довольно часто, у других его можно было видеть в крайне редких случаях.
Первая стадия формирования условной оборонительной реакции соответствует начальному периоду становления рефлекса, т. е. периоду неустойчивых ответов, появляющихся с более длительным латентным периодом, чем на последующих этапах.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'