Глава II. Взаимодействие специфической и неспецифической афферентации и его роль в формировании безусловных и в начальный период становления условных рефлексов
Изменения электрической активности специфических и неспецифических систем мозга при осуществлении безусловных реакций (макроэлектродное исследование)
Как хорошо известно, всякое экстеро-, интеро- или проприоцептивное раздражение, если оно достигает пороговой интенсивности, вызывает ответную деятельность организма, эффекторную реакцию. Характер и величина этого рефлекторного ответа зависят от модальности и интенсивности стимуляции.
Как было показано в исследованиях Pfluger (1853), И. М. Сеченова (1863), Sherrington (1906) и многих других, в нервной системе может, при достаточной интенсивности раздражения, наблюдаться широкая иррадиация возбуждения, которая выражается в вовлечении в реакцию многих эффекторных аппаратов. С этого периода, вплоть до 20-х годов нашего столетия, полагали, что пороговое раздражение вызывает всегда локальную реакцию эффектора и, следовательно, распространение возбуждения в пределах одной какой-то рефлекторной дуги.
Однако оказалось, что каждой локальной эффекторной реакции всегда сопутствует широкая иррадиация возбуждения в центральной нервной системе.
Л. А. Орбели и К. И. Кунстман (1924) первыми обратили внимание, что деафферентированная конечность реагирует на всевозможные раздражения, исходящие из внешней и внутренней сред организма. На основании этих опытов Л. А. Орбели (1938) заключил, что нервные импульсы широко иррадиируют. Наблюдения Л. А. Орбели и К. И. Кунстман были подтверждены А. Н. Черневским (1935), а также А. И. Шумилиной (1949а, б, в) и другими. Генерализованные изменения нервных процессов при действии различных раздражений были обнаружены и в электрофизиологических наблюдениях (Berger, 1930; М. Н. Ливанов, 1938; С. А. Саркисов и М. Н. Ливанов, 1933; Ectors, 1936; Rheinberger, Jasper, 1937: Gerebtzoff, 1940; Derbyshire и сотр., 1936; Forbes, Morison, 1939, и др.).
Анализ механизма этих влияний (И. М. Сеченов, 1863; В. М. Бехтерев, 1896; Н. А. Рожанский, 1935; И. С. Беритов, 1937а, б; А. И. Шумилина, 1949а, б, в; Dempsey, Morison, 1942; Moruzzi, Magoun, 1949; Jasper, 1949, и др.) позволил развить точку зрения, что ретикулярная формация, которая играет большую роль в деятельности организма, являясь субстратом рефлекторных (вегетативных и соматических) функций, имеет отношение к проведению диффузного неспецифического возбуждения.
Было высказано предположение, что генерализованные изменения ЭЭГ коры, наступающие при осуществлении ориентировочных реакций, определяются влияниями, исходящими из ретикулярной формации ствола мозга (Dongier и сотр., 1957; Gastaut, 1957; Г. Гасто, 1958; Hernandez-Peon, 1957, и др.). Так ли это в действительности?
Напомним, что весь комплекс структур, через который, как предполагают, осуществляется генерализованное влияние внешней и внутренней среды организма, был назван "системой диффузных проекций" на кору головного мозга (Dempsey, Morison, 1942) или "восходящей активирующей системой" (Moruzzi, Magoun, 1949). В отличие от специфических сенсорных путей, оказывающих локальное влияние и оканчивающихся в ограниченных территориях коры, ее также называют неспецифической проекционной системой (Ajmone-Marsan, Dillworth, 1953, и др.).
В эту систему мозга включают ретикулярную формацию ствола мозга (Magoun, 1944, 1950, 1952, 1954; Rhines, Magoun, 1946; Moruzzi, Magoun, 1949), суб- и гипоталамус (Murphy, Gellhorn, 1945; Rhines, Magoun, 1946; Moruzzi, Magoun, 1949, Magoun, 1950, 1952, 1954), ядра средней линии, интраламинарную группу таламических ядер (Moruzzi, Magoun, 1949; Jasper, 1949; Austin, Jasper, 1950).
В последние годы число образований, которые по ряду признаков можно причислить к так называемым "активирующим системам", настолько увеличилось, что исследователи стоят перед перспективой включения в это понятие большинства структур мозга. Какие данные могут быть привлечены для обоснования этого предположения?
На основании анализа опытов, в которых исследовались влияния раздражения и разрушения различных образований мозга на те или иные безусловные рефлексы (Т. С. Наумова, 1963), мы пришли к заключению, что функции, которые первоначально приписывались ретикулярной формации и другим неспецифическим системам мозга, свойственны весьма широкому кругу образований, которые также обладают особенностями, присущими "активирующим" системам (формации обонятельного мозга, образования экстрапирамидной системы и прочие). Более того, эти свойства присущи и специфическим структурам (правда, в достаточно ограниченной степени). Так, например, Starzl и сотр. (1951а, б) наблюдали генерализованные эффекты при высокочастотной стимуляции специфических систем и ответы в этих системах на раздражения других модальностей. Vernier и Galambos (1957) подтвердили эти наблюдения в микроэлектродных записях активности отдельных клеток внутреннего коленчатого тела, показав, что 20% его нейронов реагируют на свет и не реагируют на звук. На возможность проведения неспецифических (для данного образования) афферентных влияний через подкорковые ядра специфических систем указывали и другие исследователи (Неrnan-dez-Peon, Hagbarth, 1955: А. Фессар, 1961; Р. А. Дуринян, 1964). Эта мысль нашла подтверждение и со стороны нейроморфологов (Nauta, Whitlock, 1954, и др.).
Хотя с морфологической точки зрения к собственно ретикулярным структурам ствола мозга относятся только ретикулярная формация ствола, гипоталамуса, субталамуса (Riley, 1943), а также (по данным Т. А. Леонтович, 1959) парафасцикулярный комплекс ядер, ретикулярные и паравентрикулярные ядра таламуса, ядра передней комиссуры, бледный шар, диагональное поле, безымянная субстанция и ряд других, в различных отделах анализаторов обнаруживаются нейроны ретикулярного типа. Так, Т. А. Леонтович (1959), Г. П. Жукова (1959) и Е. Г. Школьник-Яррос (1959) доказали, что в специфических ядрах ствола мозга и таламуса новорожденных щенят имеются нейроны, характерные для ретикулярных структур.
Таким образом, кроме тех особенностей, которые свойственны деятельности каждой мозговой формации и характеризуют ее как образование со специфическими функциями (для большого числа структур последние еще остаются неизвестными и поэтому условно называются неспецифическими), этим же образованиям присущи общие черты, которые выявляются как при их разрушениях, так и при регистрации их электрической активности.
Общим диффузным процессам отводят роль тонизирующего возбуждения, создающего условия для более тонких и точных процессов в мозге.
Значение взаимодействия общих и частных или диффузных и локальных или, наконец, тонических и фазных процессов изучено еще очень неполно и нуждается в специальном анализе в отношении как низшей, так и высшей нервной деятельности.
Особенно важно для решения вопроса о механизмах и локализации временной связи, т. е. встречи условного и безусловного возбуждений, провести анализ взаимодействия процессов возбуждения и торможения, вызванных стимулами разных модальностей на стволово-подкорковом уровне анализаторных систем, которое до настоящего времени обследовано недостаточно. Авторы изучали проблему взаимодействия специфической и неспецифической афферентации преимущественно в отношении кортикальных формаций. Даже в случае взаимосвязи специфических и неспецифических ядер таламуса ее относят за счет коры головного мозга (С. П. Нарикашвили и Э. С. Мониава, 1961).
При изучении этого вопроса необходимо было в первую очередь параллельно исследовать электрические реакции так называемых специфических и неспецифических образований мозга при действии внешних раздражений разных модальностей.
Специального сопоставления электроэнцефалографических реакций специфических и неспецифических систем ствола мозга бодрствующих собак в ответ на действие экстероцептивных раздражений до настоящего времени проведено не было. Немногочисленные работы, посвященные параллельному изучению электрических процессов специфических и неспецифических систем на стволово-подкорковом уровне бодрствующих кошек и обезьян (А. Б. Коган, 1949; С. П. Нарикашвили, 1950; Sharpless, Jasper, 1956; В. Маунткасл, 1961), показали значительное сходство изменений электрической активности разных субкортикальных и стволовых образований мозга при общих изменениях функционального состояния и дали основание говорить о том, что в реакцию на внешние раздражения вовлекается широкий круг структур мозга, однако эти авторы не сообщают, имеется ли при этом коррелятивная связь между электрическими процессами в так называемых специфических и неспецифических системах мозга.
Для освещения этих вопросов необходимо было провести изучение электрических реакций в разных специфических и неспецифических системах ствола мозга собак при воздействии экстероцептивных раздражений разных модальностей.
Изучение электрической активности мозга собак, начатое еще В. Я. Данилевским (1891), В. Е. Ларионовым (1899), Nemminski (1913, 1925) и продолженное И. И. Лаптевым (1941, 1949), О. В. Верзиловой (1955, 1958), Л. Г. Трофимовым и сотр. (1955), Г. Т. Сахиулиной (1957, 1958, 1960), Р. С. Мнухиной (1958, 1960, 1961), В. Н. Думенко (1960а, б) и другими показало, что ЭЭГ коры головного мозга этих животных, как и у прочих млекопитающих, представляют собой сложные кривые, в которых исследователи выделяют ряд частотных составляющих - от колебаний, длящихся секунды (В. Я. Данилевский, 1891; В. Е. Ларионов, 1899), до многих десятков колебаний в 1 сек (Nemminski, 1913, 1925; Р. Н. Лурье и сотр., 1956; Г. Т. Сахиулина, 1957, и др.).
Доминирующий ритм ЭЭГ любой области мозга собаки в покое значительно более высокочастотен (до 50-60 в 1 сек), чем ЭЭГ мозга других млекопитающих, таких как кошка, кролик, обезьяна, а также человек.
Электрические процессы субкортикальных и стволовых зон мозга собаки мало чем отличаются от корковых (Л. Г. Трофимов и сотр., 1958; Yoshii и сотр., 1958а). Это сходство у бодрствующих животных в покое бывает особенно отчетливо выражено, если запись электрической активности коры, так же как глубинных структур, происходит погружными электродами, регистрирующими суммарную активность ее слоев.
В наших опытах одновременное исследование электрических явлений разных звеньев слухового анализатора (кохлеарных ядер, трапециевидного тела, верхнеоливарного комплекса ядер, ядер латеральной петли, заднего двухолмия, внутреннего коленчатого тела, коры эктосильвиевой и сильвиевой извилин), двигательного анализатора (ядер нежного и клиновидного канатиков, вентрального ядра таламуса, коры сигмовидной извилины), неспецифических и других подкорково-стволовых образований (ядер ретикулярной формации продолговатого, среднего мозга, хвостатых ядер, гиппокампа и т. д.) и различных корковых и подкорковых формаций собак показало, что при изменениях функционального состояния, при осуществлении ориентировочных реакций, вызываемых болевыми, звуковыми и световыми раздражениями, происходят изменения потенциалов, однозначные для разноименных структур каждого уровня мозга. Они выражаются в записях 40-100-микронными электродами в виде двух внешне противоположных эффектов: депрессии (снижения ритма и амплитуд) и экзальтации (одновременного повышения частотно-амплитудного показателя) электрической активности. Обе реакции носят генерализованный характер, т. е. сходно выражаются в специфических и неспецифических системах мозга.
Появление той или иной электрографической реакции на световые, звуковые и электрокожные раздражения зависит от исходного функционального состояния, индивидуальных особенностей нервной системы животного, а также модальности и интенсивности применяемого стимула.
Большое значение для проявления того или другого эффекта имеет исходное функциональное состояние животного. Например, у животных, не привыкших к экспериментальной обстановке, при действии непрерывных раздражений трех модальностей (свет, звук, электрическая стимуляция кожи лап) наблюдается значительная активация электрических колебаний образований подкорково-стволового и коркового уровней. По мере привыкания животного к экспериментальной обстановке те же раздражители вызывают снижение электрической активности. Степень выраженности того и другого эффекта индивидуальна для каждого животного, в то же время она зависит от локализации регистрируемых процессов: в отведениях от стволовых структур колебания частотно-амплитудного показателя электрической активности происходят в более широких пределах, чем в отведениях от других субкортикальных формаций мозга. Противоположные реакции могут наблюдаться и в одном опыте, на фоне лишь незначительно отличающейся исходной активности. Примером этого является запись ЭГ структур коры и продолговатого мозга собаки № 8 в одном и том же опыте, при действии одного и того же звукового раздражения - тона 500 гц 39 дб над порогом слышимости человека (рис. 6). Видно, что в первом случае этот сигнал вызвал мощную активацию: увеличение амплитуд колебаний вдвое и возрастание их ритма. На фоне несколько более активного состояния, когда исходная амплитуда колебаний в некоторых областях составляла 120-150% от исходной ЭГ в предыдущем случае, звук сопровождался постепенным снижением (в 2-3 раза) амплитуд электрических колебаний и их урежением.
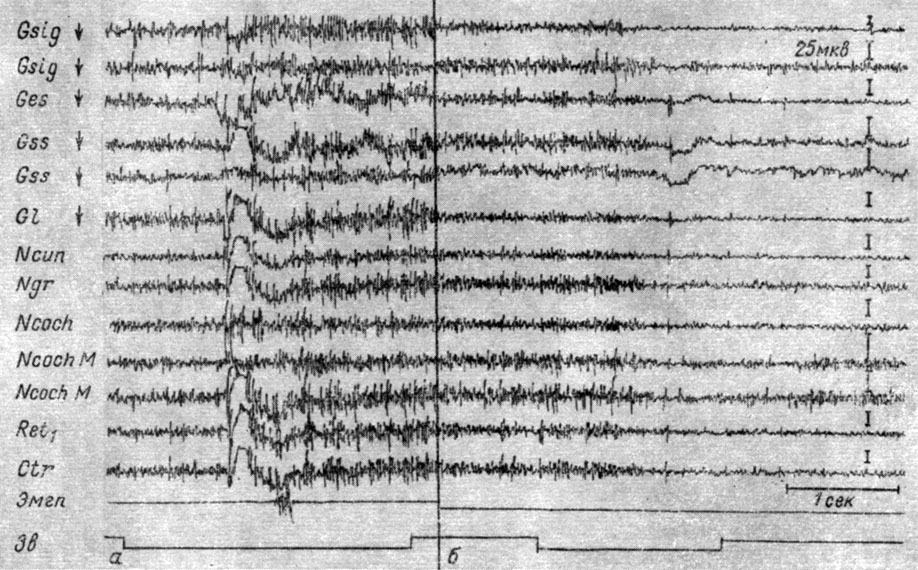
Рис. 6. Противоположные изменения ЭГ коры большого мозга и структур продолговатого мозга собаки № 8, наступающие при действии одного и того же звукового стимула (тона 500 гц 39 дб) на фоне разных исходных функциональных состояний - а и б (опыт № 14 от 26/IX 1959 г.)
Обращает внимание однонаправленность процессов в разных отделах мозга и значительное сходство их в специфических и неспецифических системах данного уровня ствола. Сходство это выступает не только в специфических и неспецифических системах ствола, но также и разных уровнях мозга - например, образованиях продолговатого мозга и коры. Это объясняется тем, что запись электрической активности коры происходила биполярным способом тонкими (60 мк) погружными электродами, регистрирующими суммарную активность слоев коры. В таких случаях, как уже указывалось, в действии экстероцептивных раздражений отчетливо тождество в изменениях ЭГ коры и подкорково-стволовых структур.
"Спонтанные" изменения ЭГ как кортикальных, так и субкортикальных формаций, а также реакций на любые внешние раздражения в большой мере зависят от индивидуальных особенностей нервной системы подопытных животных.
Характер изменений электрической активности находится в большой зависимости и от силовых свойств раздражителя. При подпороговых (в смысле вызова электрографической реакции) интенсивностях раздражения не вызывали никаких видимых изменений ЭГ. Действие раздражений, превышающих пороговые значения, обычно приводило к росту амплитуд и частот колебаний, если стимул был менее интенсивен, и к снижению этих показателей при более интенсивной стимуляции. Так, на рис. 7 показано, что у собаки № 11 действие порогового электрокожного раздражения (а) вызывает возрастание (в 1,5 раза) амплитуд колебаний электрической активности при слабом приросте их частот. В последействии от раздражения частота колебаний снижается - на фоне быстрой активности начинают регистрироваться острые редкие (12-16 в 1 сек) отклонения. Эти процессы охватывают главным образом структуры продолговатого мозга: кохлеарные ядра, гигантоклеточное ядро ретикулярной формации, средние ножки мозжечка, а также кору коронарной и сильвиевой извилин (при отведении погружными электродами). В ЭЭГ тех зон коры, которые регистрируются накладными электродами, происходит, наоборот, снижение в 2-1,5 раза амплитуд колебаний при возрастании их ритма, т. е. типичный эффект десинхронизации. При действии более сильного болевого раздражения (см. рис. 7, б) вслед за мгновенной вспышкой активности наступало резкое генерализованное ее снижение. Амплитуды колебаний упали в несколько раз, частота их понизилась на 8-10 в 1 сек. Лишь в одном из отведений от средней ножки мозжечка и отведении накладными электродами от коры коронарной извилины в меньшей степени выражено падение амплитуд колебаний, которое совпадает с возрастанием их частоты. В последействии от раздражения, длившемся 2 сек, региональные различия сглаживаются.
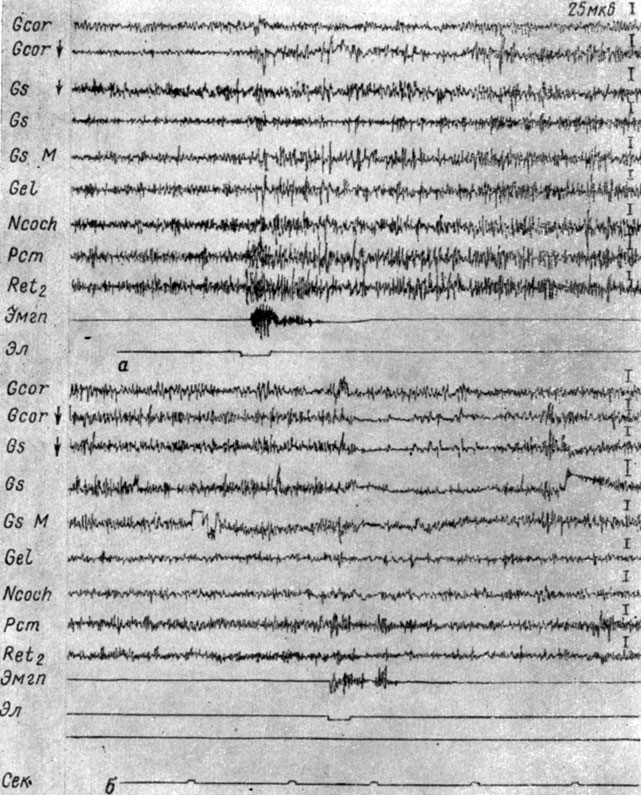
Рис. 7. Противоположные изменения ЭГ коры большого мозга и структур продолговатого мозга собаки № 11 при действии электрического тока разной интенсивности на правую переднюю лапу. а - пороговое; б - надпороговое болевое электрокожное раздражение (опыт № 10 от 6/IV 1960 г.) (по Т. С. Наумовой, 1962 а)
Как следует из только что приведенного наблюдения, эффект депрессии электрических колебаний наступает при действии более сильного болевого раздражения. Та же зависимость существует и в отношении звуковых и световых стимулов. Если они достаточно интенсивны, то вызываемый ими эффект выражается в угнетении (снижении ритма и амплитуд) электрических процессов стволово-подкоркового и кортикального уровней мозга.
Как уже говорилось, при записях потенциалов погружными электродами диаметром 40-100 мк обнаруживается идентичность изменений потенциалов коры и ствола.
Что касается глубины той или иной реакции в специфических и неспецифических системах мозга, то она может быть несколько различной, причем большие колебания активности обнаруживаются в ретикулярных и других неспецифических структурах мозга, в разных участках которых, по данным И. Н. Книпст (1961), в разные интервалы времени могут регистрироваться различные по величине местные высокоамплитудные колебания потенциала (МВКП).
Нужно отметить, что (насколько можно судить по чернилопишущей записи) та или иная реакция появляется более или менее одновременно в специфических и неспецифических структурах каждого данного уровня мозга, причем латентный период ее появления варьирует в разных экспериментальных условиях в весьма широких пределах - от 20-40 мсек до десятых долей секунды. Возможная точность измерения соответствует длительности одной полуволны, т. е. 15-20 мсек.
Одновременные и одинаковые изменения суммарной электрической активности, наступающие в специфических и неспецифических системах каждого уровня ствола мозга в ответ на действие внешних раздражений разных модальностей обнаруживаются и методом вызванных потенциалов (Galambos и сотр., 1956; Г. В. Мегун, 1958; Л. Г. Трофимов и сотр., 1958, 1959, и др.). Однако вызванные ответы, отводимые с помощью использовавшихся тонких электродов, уловить удается лишь на фоне умеренной реакции депрессии или экзальтации. На фоне же глубокой депрессии и мощной экзальтации вызванные потенциалы не обнаруживаются. Как правило, вызванные потенциалы уменьшаются на фоне депрессии ЭГ (рис. 8, а), возрастают на фоне ее экзальтации (см. рис. 8, б). Следует отметить, что световые стимулы провоцируют менее сильные сдвиги суммарной электрической активности и, видимо, поэтому в ответ на их ритмическое применение генерализованные вызванные потенциалы проявляются в более отчетливой форме. Возможно, это обусловлено и более медленным развитием ответов, вызываемых световыми вспышками.
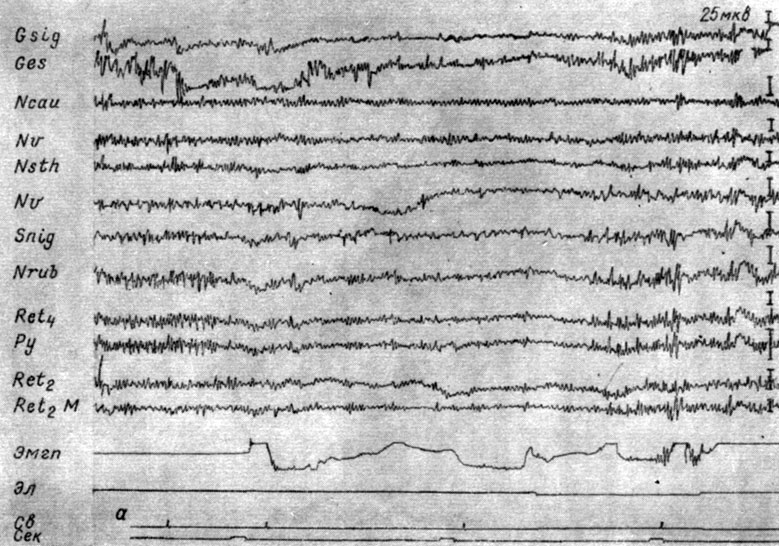
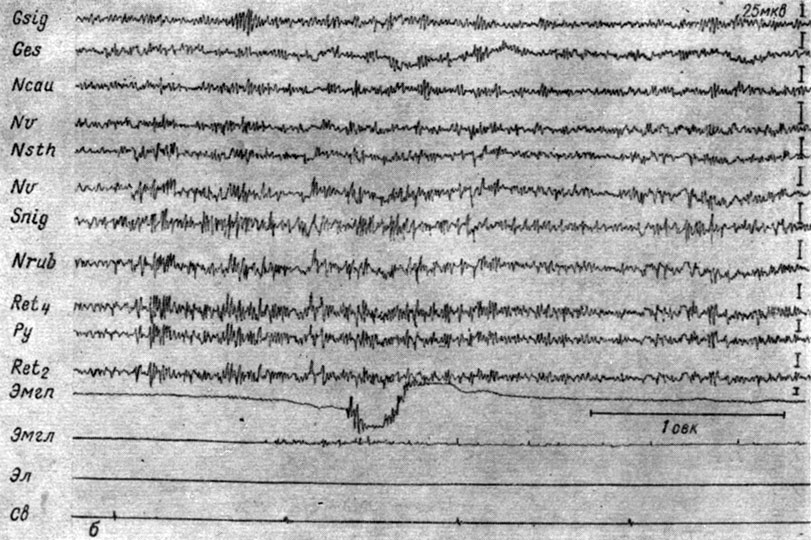
Рис. 8. Изменения величины вызванных ответов, наступающих в разных структурах мозга собаки № 15 на действие ритмических (1 в 1 сек) вспышек света (0,25 лк) в зависимости от стадии формирования на них условного оборонительного рефлекса. а - I стадия, 28-е сочетание (опыт № 93 от 14/IX 1962 г.); б - II стадия. 82-е применение (опыт № 120 от 13/III 1963 г.)
Как показали наши наблюдения, широко иррадиированные вызванные ответы регистрируются и в первых переключательных реле анализаторов, к которым данный раздражитель непосредственно не адресуется. Сказанное иллюстрирует рис. 9, на котором представлены записи ЭГ разных зон коры, сделанные погружными электродами, а также структур продолговатого мозга собаки № 8 при ритмических (10 в 1 сек) световых вспышках (0,25 лк). Можно видеть, что в ответ на действие ритмических вспышек наступает возрастание активности, отчетливо выраженное в структурах продолговатого мозга и менее четкое в коре. Первую секунду действия сигнала отчетливо заметно следование ритму раздражения в кохлеарных ядрах, в следующую секунду наступает мощная (в 2 раза по сравнению с фоном) активация как в этих ядрах, так и в ядре клиновидного канатика, а также сигмовидной, эктосильвиевой, супрасильвиевой и латеральной извилинах коры. В остальных корковых структурах, ретикулярной формации и ядре клиновидного канатика наблюдается лишь возрастание частоты колебаний без существенных изменений их амплитуд. На фоне такой активации вызванные потенциалы выделить удается лишь в ядре клиновидного канатика. Измерения латентных периодов генерализованных вызванных ответов, записанных на шлейфном осциллографе, показали, что они зависят от того, регистрируются ли ответы в системе специфических проводников данного анализатора или в других формациях, а также от уровня исследуемой структуры.
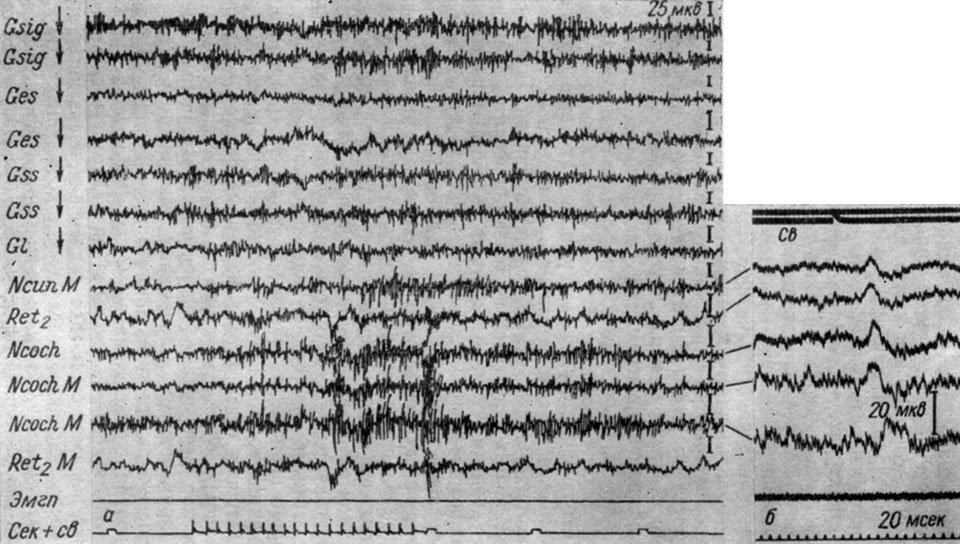
Рис. 9. Изменения ЭГ коры большого мозга и структур продолговатого мозга собаки № 8 (а) и № 4 (б) при действии прерывистых (а - 10 в 1 сек; б - 1 в сек) вспышек света (0,25 лк). а - опыт № 10 от 22/VII 1959 г.; б - опыт № 5 от 30/XII 1958 г.
Наши наблюдения, свидетельствующие о сходстве ответных реакций разных образований каждого данного уровня, обнаруженном по показателям суммарной и вызванной активности в самых различных структурах не только коркового, но и подкоркового уровня мозга, подтверждают полученные ранее данные (Galambos и сотр., 1956; Hernandez-Peon и сотр., 1956а, b; Л. Г. Трофимов и сотр., 1958; Г. В. Мегун, 1958; John и сотр., 1964), которые свидетельствуют о широкой иррадиации вызванных ответов в подкорково-стволовых образованиях мозга.
Вызванные ответы при определенных условиях, например при редких ритмах раздражения, взаимосвязанно меняются в специфических и неспецифических системах мозга. Их наличие или отсутствие зависит от того функционального состояния, на фоне которого происходит действие ритмического раздражения (Л. Г. Трофимов и сотр., 1958). Удивительно, что имеется строгая коррелятивная связь формы вызванных ответов в различных подкорковых образованиях мозга (John и сотр., 1964).
До сих пор речь шла об изменениях, вызываемых однократными применениями непрерывных или прерывистых стимулов. Если же раздражитель применяется неоднократно, то после окончания его действия возможно сохранение следовых эффектов. В отведениях корковыми накладными электродами этот эффект выражается в виде сохранения навязанных ритмов. Что касается записей, сделанных с коры или ствола погружными электродами, то он состоит в преимущественной активации той анализаторной системы, которая подвергается стимуляции. Такие изменения, наступающие в ЭГ кохлеарных ядер и слуховой коры, удалось заметить у 2 собак (№ 17, 18). Процессы, регистрируемые на следах раздражений, если последние не применяются более одного-двух десятков в опыт, через несколько дней сглаживаются, и по окончании очередных раздражений не удается обнаружить региональных различий ЭГ.
Таким образом, любая реакция на экстероцептивные стимулы является реакцией, в которую в той или иной степени, но вполне отчетливо, вовлекаются все структуры мозга. Это выступает как в изменениях суммарной электрической активности, так и в эффектах вызванных потенциалов.
Факты, свидетельствующие о сходстве изменений электрической активности специфических систем и ретикулярной формации ствола мозга, как нам представляется, говорят о том, что ретикулярная формация и другие неспецифические системы не являются единственными, через которые осуществляется диффузная афферентация.
Имеющиеся материалы дают основание думать, что диффузная коллатеральная информация идет в кору по всему стволу мозга. Иными словами, неспецифическая рассеянная афферентация коры осуществляется не только ретикулярной и другими неспецифическими формациями (Moruzzi, Magoun, 1949; Magoun, 1952, и др.), но также и специфическими системами мозга.
По-видимому, ретикулярная формация ствола мозга является местом лишь наибольшей концентрации неспецифических афферентов, поэтому при ее раздражении проще получить неспецифические ответы коры, в ней легче зарегистрировать ответы на стимуляцию различных рецепторов. Таким образом, диффузные процессы или неспецифические функции не могут являться прерогативой только ретикулярных структур мозга, - они есть общее в физиологии всех его образований.
Накопленный современной нейрофизиологией и нейроморфологией опыт подтверждает высказанное Л. А. Орбели (1938) предположение, что всякое раздражение вызывает эффект во всей центральной нервной системе.
Само собой разумеется, что развиваемый нами взгляд отнюдь не отрицает значимости локальных, более срочных процессов, которые лежат в основе наиболее дифференцированных реакций организма, а, наоборот, предполагает их еще более тесную связь, которая осуществляется уже на уровне самих анализаторных систем, начиная с их первых переключательных реле.
Очевидно, надо отказаться от неправильного представления о том, что только через ретикулярные структуры осуществляется коллатеральная информация коры головного мозга. На основании этого представления можно было бы ожидать значительные различия в реакциях специфических и неспецифических систем на действие безусловных стимулов. Однако нам удалось наблюдать лишь однонаправленные изменения частотной и амплитудой характеристик электрического процесса: или нарастание частоты и величины колебаний электрической активности (реакция экзальтации) или их одновременное снижение (реакция депрессии). Обе реакции возникают на действие любых внешних раздражений, и появление того или иного эффекта зависит от модальности и интенсивности действующего экстероцептивного стимула, индивидуальных особенностей и функционального состояния нервной системы животного. На уровне продолговатого мозга корреляция процессов обнаружена в кохлеарных ядрах, верхнем оливарном комплексе ядер, трапециевидном теле, ядрах нежного и клиновидного канатика, ядрах тройничного нерва, ядрах ретикулярной формации - nucl. medullae oblongatae centralis, nucl. gigantocellularis, nucl. parvocellularis, средней и нижней ножках мозжечка и др.; в образованиях среднего мозга: ядрах латеральной петли, внутреннем коленчатом теле, переднем и заднем двухолмии, ядрах ретикулярной формации среднего мозга - nucl. ruber, substantia nigra, nucl. cuneiformis, substantia grisea centralis; nucl. subcuneiformis; структурах межуточного мозга; вентральном и ретикулярном ядрах таламуса, субталамическом ядре и т. д., т. е. в анализаторных и, условно говоря, межанализаторных (или специфических и неспецифических) формациях каждого уровня мозга.
Без сомнения, нельзя исключить мысль, что в механизме изменений электрических процессов специфических и неспецифических систем мозга на подкорково-стволовом уровне при осуществлении всякого рода ориентировочных реакций играет большую роль взаимодействие этих систем на уровне коры (Dempsey, Morison, 1942; Bremer, Bonnet, 1950; Moruzzi, Magoun, 1949; Jasper, Ajmone-Marsan, 1952; Gellhorn и сотр., 1954; Jasper и сотр., 1955; Hernandes-Peon и сотр., 1956а, b; Gauthier и сотр., 1956; Stoupel, 1958; Dumont, Dell, 1960; Bremer, Stoupel, 1958; 1959a, b; Dumont и сотр., 1959; С. П. Нарикашвили и Э. С. Мониава, 1961, и др.) и влияние последней на субкортикальные структуры (Л. Г. Воронин и Е. Н. Соколов, 1955, 1958, 1960). Как известно, эти влияния осуществляются как на специфические (Ogden, 1960), так и на неспецифические системы мозга (Livingston и сотр., 1953; French и сотр., 1955; Дж. Д. Френч, 1958; С. П. Нарикашвили, 1960, и др.), причем, по мнению Дж. Вада (1958), последние в большей мере находятся под влиянием коры, чем первые.
Однако, так как в специфических системах реакция основной электрической активности на все виды раздражения чаще всего бывает такой же, как в ретикулярной формации, и это сходство выступает как при изменениях функционального состояния, так и при осуществлении различных ориентировочных реакций (на звук, свет, болевые раздражения), то эти данные дают основание говорить о действии раздражителей не только на специфические системы соответствующей модальности и неспецифические (ретикулярные) образования мозга, но также (и это особенно важно подчеркнуть) на специфические системы других анализаторов.
Следует указать, что само проведение ретикулярных влияний уже определяет широкий охват мозговых структур диффузным возбуждением (выделено, по крайней мере, 6 возможных путей проведения ретикулярного возбуждения к коре, в числе которых следует отметить специфические системы анализаторов). Но вполне вероятно, что диффузное распространение возбуждения может осуществляться и за счет прямых связей анализаторов, которые достаточно широко представлены в мозге, а также за счет прямых связей анализаторов с теми или иными формациями мозга (С. Б. Дзугаева, 1965; Ю. Б. Бирючков, 1963).
Наши наблюдения, полученные в хронических опытах, ставят вопрос о возможности конвергенции возбуждений разного происхождения на уровне специфических систем ствола мозга.
Для того чтобы проанализировать механизмы возможного взаимодействия нервных процессов на уровне специфических систем мозгового ствола, было предпринято микроэлектродное исследование.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'