Глава I. Методика исследования
В работе было использовано 30 собак, из которых у 4 исследовалась электрическая активность только коры головного мозга и у остальных изучались электрические процессы как коры большого мозга, так и различных подкорковых и стволовых ядер (всего около 40 образований).
Для отведения электрических потенциалов в разные образования мозга подопытных собак вводились электроды, которые представляли либо нихромовые (диаметром 40-100 мк), либо серебряные (диаметром 300 мк), надежно изолированные проволочки, соединенные с коммутационным устройством ("пробкой"), укрепляемым на черепе. Отведение ЭЭГ подкорково-стволовых образований головного мозга происходило с помощью нихромовых электродов, отведение ЭЭГ коры - с помощью серебряных электродов.
До последнего времени в электрофизиологической литературе существуют значительные разногласия по поводу выбора способа отведения электрической активности. Эти разногласия касаются как физиологических, так и клинических исследований (М. Н. Ливанов, 1938; А. Б. Коган, 1949; Л. А. Новикова и сотр., 1952; В. А. Адамович, 1957, и др.).
На основании многолетнего изучения электрических процессов мозга собак и кроликов в хронических и острых экспериментах, вслед за А. Б. Коганом (1949), В. А. Адамовичем (1956) и др., мы пришли к выводу о том, что в каждом случае выбор способа отведения должен диктоваться особенностями эксперимента.
Что касается регистрации процессов с глубинных структур мозга, то наши наблюдения свидетельствуют о целесообразности использования биполярного метода отведения. На рис. 1 представлены изменения электрической активности корковых и стволовых структур мозга собаки № 12 в стадии концентрации рефлекса, когда наиболее выражены локальные сдвиги ЭГ. При сопоставлении биполярных и униполярных записей ЭГ бульбарных структур мозга и коры видно значительное сходство записей электрической активности, сделанных униполярным способом (верхние 3 кривые). В этих записях наблюдаются медленные отклонения, перемежающиеся с периодами сниженной электрической активности. В это же время в записях, сделанных биполярным способом, обнаруживается значительное возрастание ритма и амплитуд электрических колебаний кохлеарных ядер, менее выраженное их возрастание в ретикулярной формации продолговатого мозга и снижение активности в ядрах нежного и клиновидного канатиков.
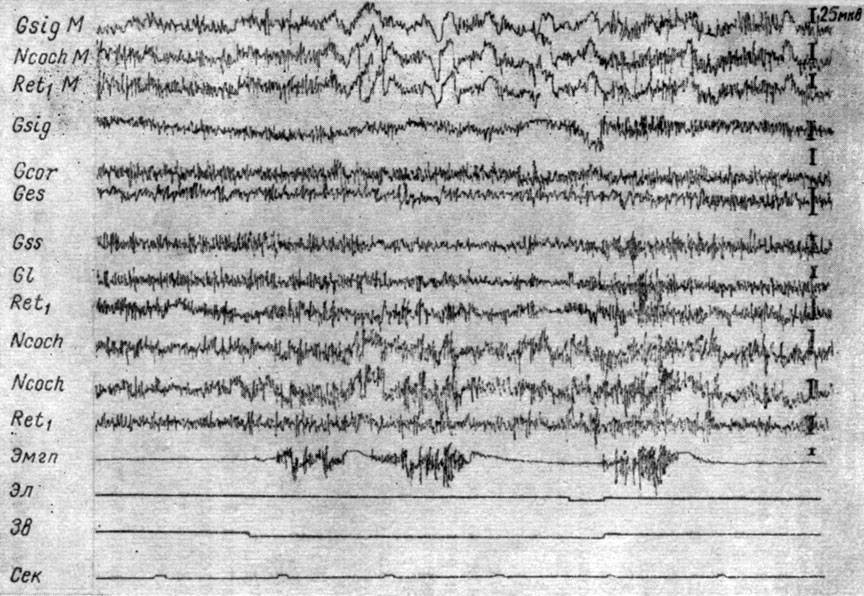
Рис. 1. Сопоставление монополярного (три верхние кривые) и биполярного способов (остальные кривые) регистрации ЭГ при осуществлении оборонительного условного рефлекса в стадии III у собаки № 12 (63-е сочетание тона 500 гц 48 дб и болевого раздражения правой передней лапы; опыт 31 от 9/II 1959 г.)
Приведенные данные показывают, что выявление локальных стволовых процессов, сопровождающих высшую нервную деятельность собак, возможно лишь при биполярном способе регистрации электрической активности.
В этом отношении следует присоединиться к мнению А. Б. Когана, который в 1949 г. писал, что "суммарный характер потенциалов, регистрируемых при "униполярном" отведении, заставил нас отказаться от этого способа, чреватого ошибками, в случае его использования для целей локализации" (стр. 46). Большое сходство записей электрической активности корковых структур при униполярном и биполярном способах отведения подтверждает мнение А. Б. Когана (1949) о том, что при регистрации униполярным способом ЭЭГ коры мозга более вероятна запись локальных процессов, чем при регистрации ЭЭГ глубоких структур.
Так как биполярный способ регистрации при малом межэлектродном расстоянии дает более точное представление о локальных электрических процессах (А. Б. Коган, 1949; В. С. Русинов, 1951; Д. С. Воронцов, 1960; Galambos и сотр., 1956; Galambos, Sheatz, 1960; Yoshii и сотр., 1958а; Н. Иошии и сотр., 1958б; Г. Т. Сахиулина, 1958; Я. Буреш и О. Бурешова, 1960; Malliani и сотр., 1963; Galvet и сотр., 1964, а также данные настоящей работы), при регистрации процессов погружными электродами использовался преимущественно этот способ. При биполярном способе отведения погружными нихромовыми электродами межэлектродное расстояние не превышало 1,5-2 мм, при записи электрических потенциалов серебряными накладными электродами оно не выходило за пределы 4-5 мм. Процессы, полученные при биполярном способе регистрации, сопоставлялись с записями ЭГ, сделанными униполярным способом.
При исследовании динамики параметров вызванных ответов применялось униполярное отведение в 2 вариантах - в первом индифферентный электрод находился на носовых костях, во втором индифферентный электрод был вынесен за пределы активной зоны (по А. И. Ройтбаку, 1958).
Погружные нихромовые электроды заключались в стеклянные трубочки, а затем попарно соединялись кольцами кембрика и склеивались полистероловым лаком (растворенным в бензоле) или оргстеклом, растворенным в дихлорэтане. Общий диаметр сдвоенного электрода не превышал 0,3-0,4 мм. Электрод выступал из стеклянной трубочки на 0,5-1 мм. Общая его длина определялась глубиной погружения.
Вживление электродов в мозг проводилось в асептических условиях под тиопенталовым (5%) внутривенным наркозом. После освобождения кости черепа на нем фиксировалось коммутационное устройство "пробка".
Для регистрации ЭЭГ структур коры накладными электродами в кости просверливались отверстия до lamina vitra, в которые вставлялись спиральки отводящих серебряных электродов, идущие от "пробки" (методика Р. Н. Лурье и Л. Г. Трофимова, 1956). Отведение ЭЭГ корковых структур у большинства животных происходило накладными электродами. У 7 собак электрические процессы коры регистрировались и с помощью погружных электродов. Погружение осуществлялось таким образом, что нижний электрод входил в нижние слои коры, а верхний располагался поверхностно - в слоях I, II (рис. 2). При таком способе отводилась суммарная разность потенциалов поперечника коры (например, слоев I-IV, I-V, II-VI и т. п.). Перед их погружением производилось просверливание отверстия в кости черепа и прокол твердой мозговой оболочки.
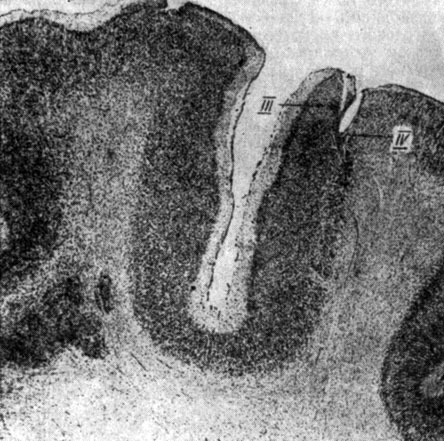
Рис. 2. Положение погружных отводящих электродов в структурах коры головного мозга собаки № 18 (слой III и IV поля Т3 - gyr. ectosylvius medius)
Так как мы располагали собаками самых различных размеров, воспользоваться стереотаксическими приборами для погружения электродов в субкортикальные отделы мозга не представлялось возможным, поэтому пришлось использовать разные способы введения электродов, как известные из литературы, так и разработанные нами специально, которые давали бы надежные результаты в смысле точности попадания отводящих электродов в соответствующие образования мозга.
Для попадания в ядра нежного и клиновидного канатиков, а также в нижнее центральное ядро ретикулярной формации в затылочной кости проделывалось отверстие таким образом, чтобы оно находилось над задним краем мозжечка. После прокола твердой мозговой оболочки электроды вводились в ствол мозга на глубину, соответствующую положению того или иного ядра.
Если электроды погружались в кохлеарные ядра, трапециевидное тело, верхнюю оливу, гигантоклеточное и мелкоклеточное ядра ретикулярной формации продолговатого мозга, среднюю ножку мозжечка и ядра тройничного нерва, это производилось через боковую поверхность продолговатого мозга после предварительного смещения вверх нижнего края flocculus мозжечка. Для осуществления этого подхода во впадине затылочной кости между затылочным мыщелком и яремным отростком просверливалось отверстие диаметром 5 мм. Этот подход к мозгу является довольно сложным, так как почти на том же уровне в твердой мозговой оболочке проходит сагиттальный синус, ранение которого чревато тяжелыми последствиями. Кроме того, при подходе к стволу приходится нарушать целость сосудистых сплетений продолговатого мозга, что связано со значительным кровотечением.
Одновременно в продолговатый мозг вводилось от 10 до 16 электродов, т. е. по 5-8 пар.
Погружение электродов в структуры среднего мозга у ряда собак производилось по способу Н. Н. Любимова и Л. Г. Трофимова (1958), который состоит в том, что после снятия верхней половины черепной крышки вводятся электроды в соответствующие образования ствола мозга по предварительным расчетам под контролем зрения. После погружения электродов провода, идущие к "пробке", укладывались строго по намету мозжечка. Более подробное описание этого способа вживления можно найти в статье Н. Н. Любимова и Л. Г. Трофимова (1958). Вводилось обычно до 10-12 электродов, т. е. до 5-6 пар.
Введение электродов в структуры среднего, межуточного и переднего мозга происходило также через кору. Такое погружение осуществлялось двумя способами.
Первый способ представлял введение каждой пары электродов в глубь мозга через маленькие отверстия, просверленные в черепе. Глубина погружения, а также положение отверстия для погружения выбирались в соответствии с координатами мозга собаки, разработанными Т. А. Леонтович и Т. А. Меринг (см. О. С. Адрианов и Т. А. Меринг, 1959).
При втором способе подкорковые электроды, предварительно смонтированные в общей стиракриловой заливке, погружались в мозг одновременно. Погружение пучка этих электродов происходило после предварительного обнажения поверхности мозга, по площади, равной площади стиракрилового основания, в котором крепятся электроды. Предварительные расчеты положения и глубины погружения делались также на основании координатов мозга собаки по данным Т. А. Леонтович и Т. А. Меринг (О. С. Адрианов и Т. А. Меринг, 1959). После введения этого пучка электродов в мозг (одновременно вводилось до 10 пар электродов) производилось прикрепление основания описанной системы к черепу с помощью жидкого стиракрила.
"Пробка" и все электроды крепились с помощью винтов и (или) стиракрила.
Нельзя не отметить, что все описанные способы вживления электродов имеют весьма существенный недостаток, который состоит в том, что во всех случаях погружение электродов связано со значительными нарушениями целости мозговой ткани, особенно в случаях повреждения сосудов. Если при погружении электродов повреждаются крупные сосуды ствола, это сопровождается кровоизлияниями, подъемом внутричерепного давления, пролапсом и т. д., о которых исследователи обычно не сообщают. Естественно, что чем больше вводится электродов, тем вероятнее получить осложнения во время и после операций.
Что касается мозговой ткани, через которую проходят кончики электродов, то в ее клеточной структуре существенных изменений не происходит. В этом отношении можно согласиться с Delgado (1955) и М. М. Александровской (1962), которые не обнаружили существенных изменений структуры мозга по ходу электродов. Но, как показала М. М. Александровская, у кончика электрода, а по нашим наблюдениям иногда также и по ходу всего электрода наступает глиальная реакция. В остальном окружающая структура остается неизменной.
Как показали наши наблюдения сопоставление процессов в кортикальных и субкортикальных структурах мозга правомочно проводить только в том случае, когда их отведение осуществляется одинаковым способом, например биполярным, погружными электродами одного диаметра с одинаковым межэлектродным расстоянием. Наши данные свидетельствуют о том, что сопоставление электрических процессов, регистрируемых погружными электродами с глубинных структур мозга и накладными (внутричерепными) электродами с коры, нередко приводит к совершенно неверным выводам.
Так как введение множества погружных электродов влияет отрицательно на состояние нервной системы подопытных животных, у большинства собак отведение ЭЭГ коры осуществлялось с помощью внутричерепных (накладных) электродов, однако в этом случае сопоставление процессов коры и подкорковых структур проводилось с учетом особенностей отведения.
Как правило, работа с подопытными животными начиналась через 2-3 недели, однако хорошие записи электрической активности погружными электродами получаются не ранее, чем через месяц, а иногда и 1,5-2 месяца после операции. В течение нескольких дней обследовалась "спонтанная" активность мозга, а также ее изменение при экстероцептивных раздражениях, затем начиналась выработка условных оборонительных рефлексов. У большинства собак она происходила по методике В. П. Протопопова (1909, 1931) и у 5 животных - по способу В. П. Петропавловского (1934).
Отводящими и раздражающими электродами лапы служили серебряные чашечки диаметром 5 мм с помещенными в них тампончиками ваты, смоченными физиологическим раствором. Электроды фиксировались эластичным бинтом на предварительно выбритой поверхности кожи правой передней лапы животного (раздражающие на нижней трети голени, отводящие - над сгибателями верхней трети голени). Расстояние между электродами варьировало от 3 до 8 см.
В качестве условных звуковых сигналов использовались тоновые раздражения 500 гц интенсивностью 30-50 дб над порогом слышимости человека, тоны 500 гц 60-100 дб и щелчки интенсивностью 56 дб, прерываемые в ритме 2 в 1 сек, 10 в 1 сек. В качестве дифференцировочных раздражителей употреблялись звуки, сходные по интенсивности с положительными сигналами, но другой частоты: тон 300 гц 30-50 дб; щелчки интенсивностью 56 дб, следующие в ритме 1 в 1 сек и 20 в 1 сек.
Применявшиеся тоновые стимулы были подпороговыми, пороговыми и надпороговыми в смысле вызова электрографической реакции у собак. Выработка условных рефлексов осуществлялась и на прерывистые световые раздражения (2 в 1 сек и 10 в 1 сек), исходящие от светового экрана, расположенного в 1 м от головы собаки. Каждая вспышка на уровне глаз животного достигала 0,25 лк. Дифференцировкой к ним служили те же световые раздражения, прерываемые в ритме 1 в 1 сек и 20 в 1 сек.
Анализ вызванных ответов в первичной проекционной зоне слуха осуществлялся при включении условнорефлекторного звукового сигнала (тона) или щелчков, следующих в ритме 2 в 1 сек (положительный рефлекс) и 1 в 1 сек (отрицательный). Редкий ритм условного стимула был избран в связи с тем, что в хронических условиях за период 500 мсек обычно завершается процесс восстановления исходного функционального состояния (В. Г. Скребицкий, 1960, 1962), и, следовательно, при ритме стимуляции 2 в 1 сек и 1 в 1 сек каждое последующее раздражение будет падать на нервную систему, когда цикл возбудимости от предшествующего раздражения оказывается завершенным.
Угашение вызванных ответов и других электрических реакций до выработки рефлекса не проводилось, они лишь обследовались в течение 4-5 опытов.
Период изолированного действия условного сигнала составлял 2-3 сек, в конце которых наносилось пороговое электрокожное раздражение на правую переднюю лапу, длительность которого не превышала 1 сек.
Как показали наши наблюдения, применение множества сочетаний в опытный день по оборонительной методике у животных с введенными в мозг электродами ведет к срывам высшей нервной деятельности. В связи с этим количество сочетаний в опыте было ограничено до 2-3.
Записи ЭЭГ проводились 8- и 16-канальными чернилопишущими электроэнцефалографами. Чернильная запись производилась на бумажной ленте, скорость движения которой составляла 60 мм в 1 сек. Регистрация вызванных потенциалов осуществлялась на 9-шлейфном осциллографе. Запись производилась на осциллографической бумаге со скоростью 120-200 мм/сек и более.
Морфологический контроль положения погружных отводящих электродов проводился на серийных парафиновых срезах мозга подопытных животных, окрашенных по методу Ниссля. В табл. I представлены сведения о локализации корковых и подкорковых погружных электродов у 16 собак, положение которых определено микроскопически. На рис. 3, 4, 5 представлены микрофотографии срезов мозгов 3 собак, на которых видны следы кончиков отводящих электродов.
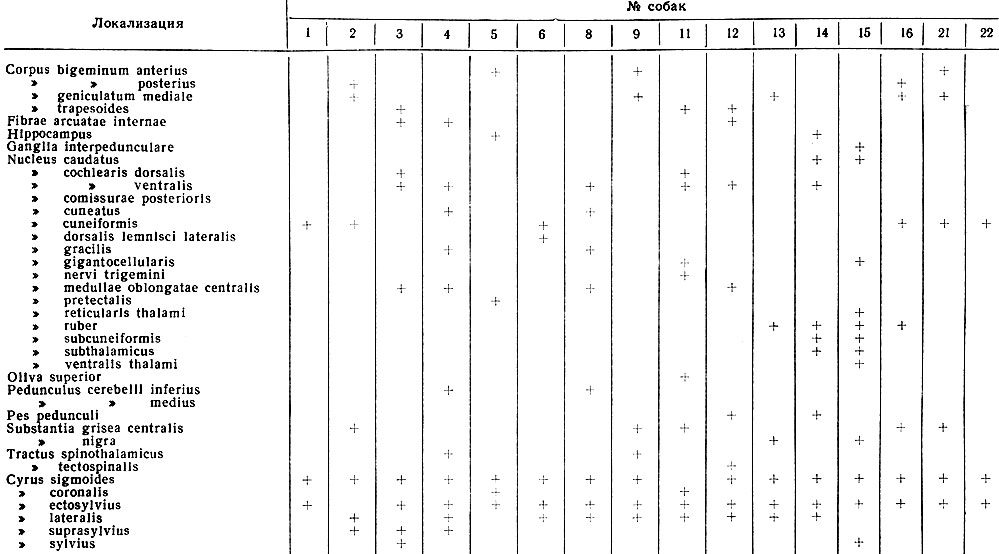
Таблица I. Локализация отводящих электродов в корковых и подкорковых структурах мозга собак
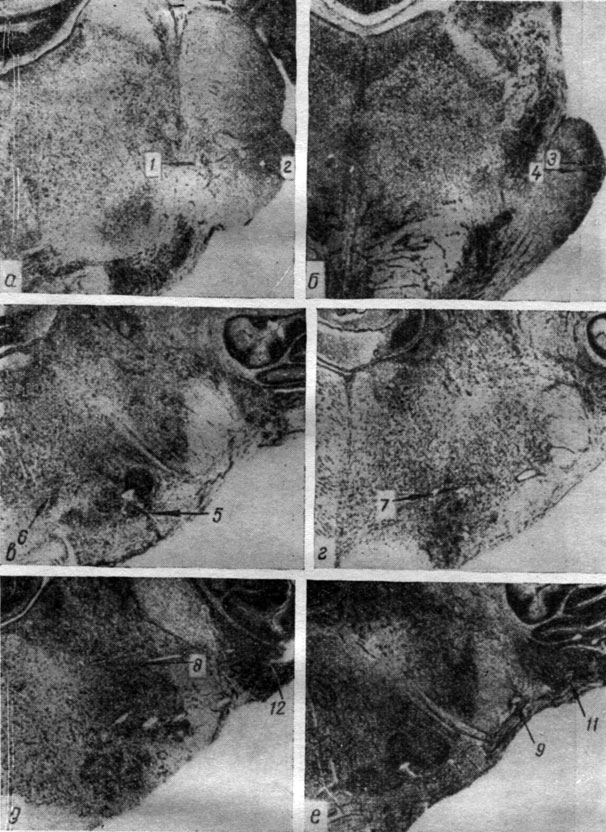
Рис. 3. Положение отводящих электродов в ядрах продолговатого мозга собаки № 11. Ув. 7:1. Номера отводящих электродов: 1 - pedunculus cerebelli medius (3а); 2, 9 - n. trigeminus (3а, 3е); 3, 4 - pedunculus cerebelli medius (3б); 5 - oliva superior (3в); 6, 7 - nucl. gigantocellularis (3в, 3г); 8 - nucl. n. trigemini (3д); 11 - nucl. cochlearis ventralis (3e); 12 - nucl. cochlearis dorsalis (3д)
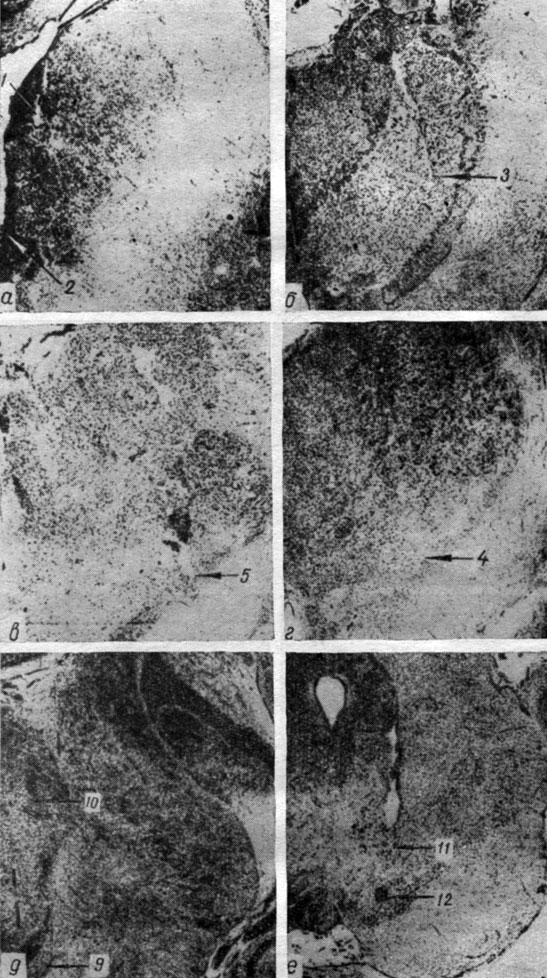
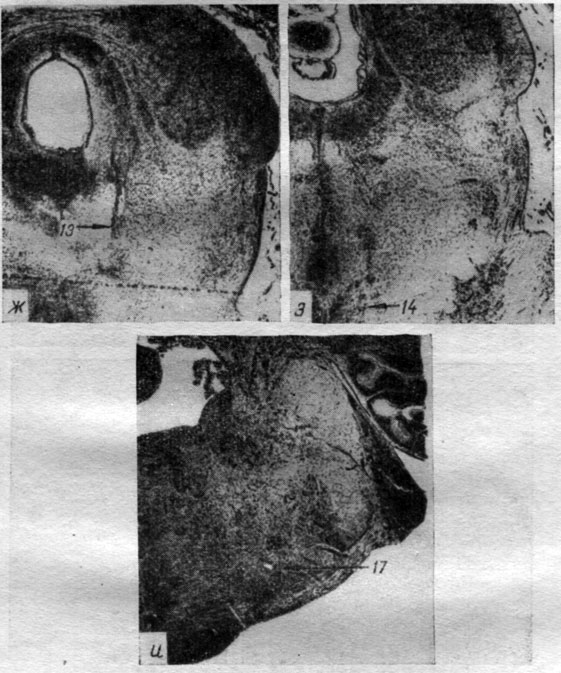
Рис. 4. Положение отводящих электродов в подкорково-стволовых структурах мозга собаки № 15. Ув. 7:1. Номера отводящих электродов: 1, 2 - nucl. caudatus (4а); 3 - nucl. ventralis thalami, pars arcuata (4б); 4 - nucl. reticularis (4г); 5 - nucl. subthalamicus (4в); 9, 12 - substantia nigra (4д, 4е); 10 - nucl. comissurae posterioris (4д); 11 - nucl. ruber (4e); 13 - nucl. subcuneiformis (4ж); 14 - nucl. interpedunculare (4з); 17 - nucl. gigantocellularis (4и)
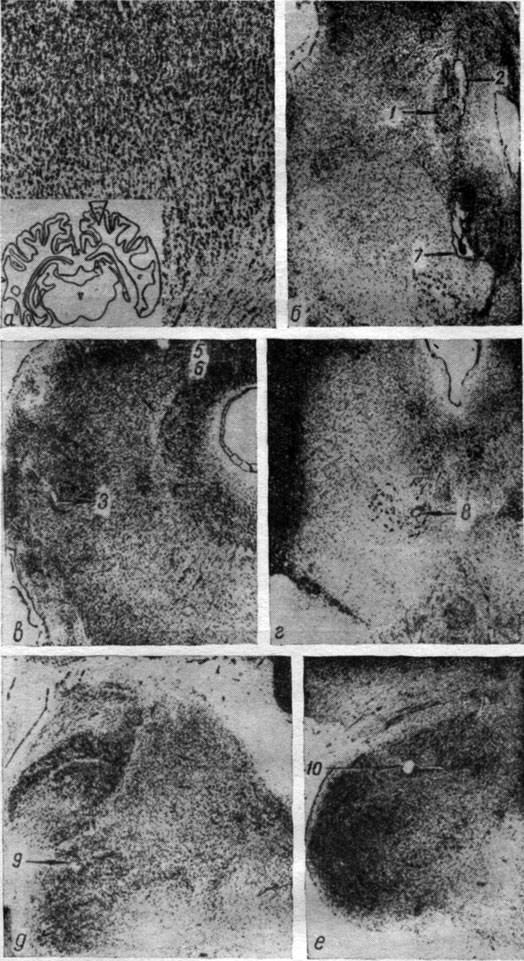
Рис. 5. Окончание отводящих электродов в структурах среднего мозга собаки № 16. Номера отводящих электродов: 1, 2 - substantia grisea centralis (5б); 3 - nucl. cuneiformis (5в); 5, 6 - corp. bigeminum posterius. (5в); 7 - fasc. tecto-spinalis (5б); 8 - nucl. ruber (5г); 9, 10 - corp. geniculatum mediale (5д, e). a - структура коры полушария, поднимавшегося во время операции. Ув. 7:1
У 10 собак расположение электродов проверено макроскопически. Контроль положения корковых накладных электродов у всех собак делался макроскопически.
Наименование корковых и субкортикально-стволовых образований дается по номенклатуре "Атласа мозга собаки" О. С. Адрианова и Т. А. Меринг (1959). Исключение составляют ядра ретикулярной формации ствола мозга, которые даны по номенклатуре Olszewski, Baxter (1954).
Несколько слов в отношении терминологии изменений ЭГ, регистрируемых тонкими погружными электродами, которые можно охарактеризовать как субмакроэлектроды.
Увеличение амплитуд и частот колебаний обозначается как экзальтация, усиление, возрастание электрической активности, которые употребляются как синонимы. Электрографическая картина, выражающаяся в снижении ритма и амплитуд колебаний, условно обозначается как депрессия, угнетение, снижение, уменьшение электрических процессов. Чтобы отличить эту реакцию от реакции, регистрируемой 300-микронными корковыми накладными электродами и выражающейся в снижении амплитуд колебаний и одновременном возрастании их частот, последняя называется эффектом десинхронизации.
Наблюдаемые изменения суммарной электрической активности были подвергнуты следующему анализу. В связи с тем, что изменения электрических потенциалов, регистрируемые с помощью погружных макроэлектродов (40-100 мк), выражаются, как правило, в однонаправленных сдвигах частотного и амплитудного показателя активности (см. след, раздел), мы сочли возможным подвергнуть исследованию один из этих показателей, а именно параметр амплитудной характеристики, колебания которого были наиболее яркими. Были проведены вычисления амплитудного показателя фоновой активности, амплитудного показателя активности в период максимально выраженного изменения ее на фоне действия условного сигнала, а также его последействия. Затем вычислялось процентное отношение реакционной и постреакционной амплитуды колебаний к ее фоновым значениям.
Полученные данные представлены в виде графиков, на которых по оси абсцисс отложены номера сочетаний, а по оси ординат - прирост (или уменьшение) амплитудного показателя электрической активности (в % по отношению к фону). В тех случаях, когда этот показатель превышал 100%, это значило, что в период действия условного сигнала и осуществления условного рефлекса (или его последействия) наступала экзальтация электрической активности. Тогда, когда тот же показатель был ниже 100%, это свидетельствовало о депрессии электрических процессов.
Мы отдаем себе отчет в том, что такой анализ дает весьма одностороннюю информацию о характере изменений электрической активности мозга, однако мы вынуждены были к нему прибегнуть в связи с тем, чтобы дать представление о динамике изменений хотя бы одного параметра электроэнцефалограммы в течение всего периода формирования условного оборонительного рефлекса. Изменения суммарной электрической активности при формировании рефлекса очень демонстративны, и представление о закономерных изменениях ЭГ на уровне мозговых структур на разных этапах становления условного оборонительного рефлекса у собак и их длительности могут дать приводимые графики. Естественно, что более глубокий анализ процессов на уровне каждой формации мозга с привлечением электронновычислительной техники бесспорно даст возможность глубже проникнуть в природу физиологических явлений при становлении рефлексов, однако сейчас приходится лишь мечтать о динамическом поликорреляционном анализе, т. е. об одновременном корреляционном анализе между несколькими процессами (10-15 как в нашем случае), сопоставляемыми за период в несколько месяцев выработки рефлекса.
Анализ вызванных ответов в первичной проекционной зоне слуха проводился путем вычисления общей величины вызванного потенциала на включение звуковых сигналов пороговой интенсивности, а также амплитудой характеристики и длительности трех первых его компонент (позитивного, негативного и второго позитивного) на каждый из 6 щелчков, следовавших в ритме 2 в 1 сек и служащих сигналом условной оборонительной реакции.
Вычислялся также латентный период начала вызванного ответа, времени до максимума первой позитивной и максимума негативной фаз ответа на каждый из тех же 6 щелчков. По этим данным строились графики, которые составлены таким образом, что по оси абсцисс отложены порядковые номера сочетаний условного сигнала с безусловным, а по оси ординат абсолютные значения амплитуд, латентного периода или длительности каждого компонента ответа на первый (непрерывная линия), третий (пунктирная линия) и четвертый (линия точек) щелчки из серии их, служащей сигналом рефлекса.
Отсутствие ответа или неясная его выраженность условно представлены на графиках как нулевая величина амплитуды или длительности.
Динамика амплитуды, латентного периода и длительности вызванных ответов представлена по показателям отведения ЭЭГ от полей Т3, PJ2 по карте О. С. Адрианова и Т. А. Меринг (1959).
Методика микроэлектродных опытов изложена в главе II.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'