Реакции клеток ядра клиновидного канатика на экстероцептивные раздражения (микроэлектродное исследование)
Обследование электрических процессов в специфических и неспецифических системах ствола мозга позволило предполагать, что проведение неспецифического возбуждения может осуществляться не только через ретикулярную формацию ствола мозга, но также через другие формации и в том числе через специфические системы (начиная с их первых реле) смежных анализаторов. Наши данные дают основание предположить, что конвергенция импульсов возможна во всех образованиях мозга, в том числе и тех, которые относятся к специфическим системам.
Изучение вопроса взаимодействия афферентаций разных модальностей в специфических системах ствола мозга не получило до сих пор достаточного освещения в литературе. Вместе с тем, его решение имеет принципиальное значение для понимания механизмов замыкания временной связи, т. е. для понимания того, может ли взаимодействие анализаторов осуществляться на разных уровнях самих анализаторов. Поэтому мы сочли необходимым обследовать процессы конвергенции импульсов разных модальностей на клетках одного из стволовых образований специфической системы. Таким образованием было избрано первичное переключательное реле кожно-двигательного анализатора - ядро клиновидного канатика.
Имеется достаточно данных о клеточной композиции и клеточных реакциях этого ядра на соматосенсорные стимулы. Известно, что нейроны ядра клиновидного канатика очень многочисленны, состоят из типичных чувствующих клеток, аксоны которых имеют коллатерали, обеспечивающие связь клеток между собой, а также с соседними специфическими ядрами и ретикулярной формацией (Ramon у Cajal, 1909). Кроме этих элементов, в состав ядра Бурдаха входят нейроны, которые, по данным Г. П. Жуковой и Т. А. Леонтович (1964), относятся к нейронам, составляющим основную массу клеток ретикулярной формации.
Как показали физиологические наблюдения, нейроны ядра клиновидного канатика могут быть возбуждены как одиночными пороговыми импульсами, так и их залпами в соматическом афферентном волокне. Therman (1941) считает, что в этом ядре происходит как моносинаптическая, так и полисинаптическая передача импульсов.
Спайки, регистрируемые экстрацеллюлярно с помощью микроэлектродов от нейронов ядра Бурдаха в ответ на раздражения соматических нервов, были или негативными или позитивно-негативными (Amassian, De Vito, 1954, 1957).
По частоте ответных залпов на периферическую стимуляцию нейроны ядра делят на 2 группы: клетки одной из них отвечают с частотой 50-380 в секунду, другой - с частотой 20 в секунду и меньше (Amassian, De Vito, 1954, 1957). Раздражения соматического происхождения могут вызывать как увеличение залпов спонтанной активности нейронов ядра клиновидного канатика, так и их торможение.
Изучая временные характеристики разрядов клеток, те же авторы заметили, что скрытые периоды ответов сильно варьируют, но тем не менее у одних нейронов они не превышают 2 мсек, у других находятся в пределах от 2 до 10 мсек. Колебания латентных периодов становятся меньше, когда используются сильные раздражения. Это объясняется тем, что при разных интенсивностях стимуляции возбуждение распространяется по путям разной длины, достигающим ядра непосредственно или через другие центральные структуры.
Вероятность залпов ответов, их число и частота с усилением стимуляции увеличиваются (Towe, Morse, 1962). Латентность и временные различия в ответах нейронов сокращаются также при увеличении числа стимулируемых афферентных волокон.
В литературе нет данных о характере реакций отдельных нейронов ядра клиновидного канатика на неспецифические для него раздражения: свет, звук. Есть лишь указания на то, что такие ответы получаются с трудом (Amassian, De Vito, 1957).
Мы попытались изучить клеточную активность ядра клиновидного канатика не только и не столько на соматосенсорную стимуляцию, сколько на звуковые и световые раздражения.
Эксперименты осуществлены на 10 кроликах и 12 кошках. Запись клеточной активности происходила на бодрствующих кроликах, оперированных под местной новокаиновой анестезией, на кошках на фоне и после пробуждения от нембуталового сна, а также под хлоралозным наркозом в сочетании с нембуталом и хлоралозным наркозом в сочетании с диплацином.
Нембутал (40 мг/кг) вводился внутрибрюшинно, хлоралоза (40 мг/кг) и диплацин (2 мг/кг) внутривенно.
Многообразие экспериментальных приемов использовалось для того, чтобы найти условия, при которых можно обнаружить реакции клеток ядра клиновидного канатика на неспецифические для них световые и звуковые раздражения.
Для отведения разрядов клеток использовались изолированные лаком стальные и вольфрамовые микроэлектроды диаметром 2-4 мк. Микроэлектрод укреплялся в держателе микроманипулятора, сделанного по конструкции А. М. Мелиховой (лаборатория М. Н. Ливанова), который прикреплялся к кости большого затылочного бугра либо зубным фосфат-цементом, либо винтиками. Регистрация клеточной активности, предварительно усиленной с помощью усилителя УБП1-01, производилась на двухлучевом катодном осциллографе, непрерывная съемка осуществлялась с помощью лентопротяжного устройства. Скорость движения пленки - 25 и 50 мм/сек.
В качестве экстероцептивных раздражителей использовались пороговая стимуляция кожи ипсилатеральной лапы или седалищного нерва электрическим током (1 в 1 сек), световые (одиночные и ритмические вспышки длительностью 1 мсек, освещенностью 300 лк на экране, расположенном на расстоянии 0,5 м от глаз животного) и звуковые раздражения (щелчки 60 дб и тоны 500 гц 50-70 дб). Раздражения давались по следующей программе: щелчки, одиночные и ритмические (1 в 2 сек, 1 в 1 сек, 2 в 1 сек), световые вспышки, одиночные и ритмические (1 в 2 сек, 1 в 1 сек, 2 в 1 сек), электрические раздражения кожи или седалищного нерва одиночные, тоны непрерывные (длительность 1-2 сек).
Интервалы между раздражениями разных модальностей составляли 1-2 мин. Длительность каждой стимуляции исходила из расчета дачи 10 одинаковых стимулов.
Расположение клетки контролировалось определением глубины погружения микроэлектрода от поверхности ядра.
Экспериментальные данные обрабатывались путем подсчета числа разрядов за период 100 мсек, на основании чего строились гистограммы.
Анализу подверглись реакции 100 нейронов.
Клетки ядра клиновидного канатика делятся на так называемые "молчащие" и "спонтанно" разряжающиеся.
В наших опытах было обнаружено 6 нейронов, импульсация в которых возникала лишь в ответ на соматосенсорные раздражения, и 94 "спонтанно" активных нейрона:
"Молчащие" нейроны приходили в активное состояние (разряжались) при раздражении электрическим током кожи лапы, седалищного нерва, а также при прикосновении к телу животного с латентным периодом, не превышающим 10 мсек. Изучение реакций этих нейронов на неспецифические для них световые и звуковые раздражения показало, что такие реакции у них обнаружить не удается.
Распределение "молчащих" нейронов по глубине расположения клеток представлено в табл. II.
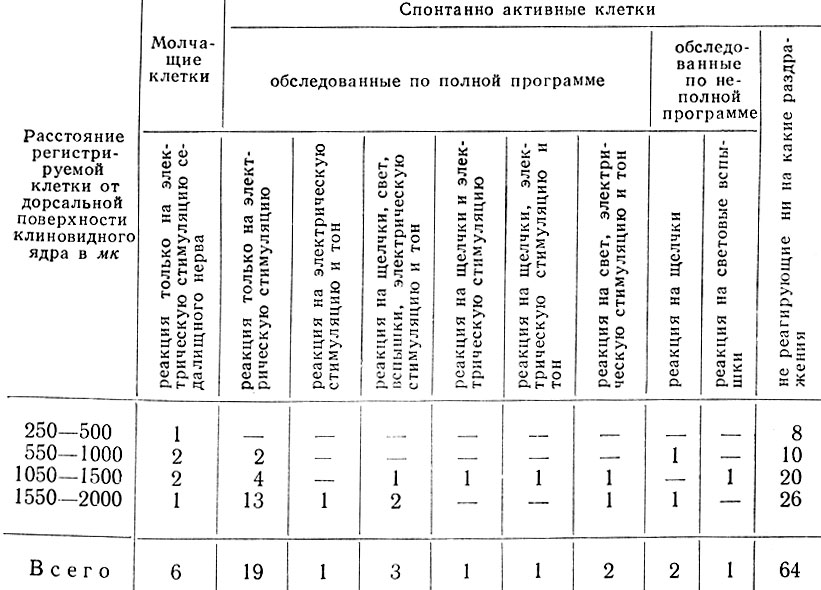
Таблица II. Особенности реакций и распределение по глубине нейронов ядра клиновидного канатика
94 спонтанно разряжающиеся клетки имели исходную частоту импульсации: 1-5 импульсов в 1 сек (15 клеток), 10-40 импульсов в 1 сек (71 клетка), свыше 50 (65-75) импульсов в 1 сек (8 нейронов).
Изучение реакций 94 спонтанно активных элементов показало, что 64 клетки, разряжавшиеся преимущественно в ритме 10-45 в 1 сек, не реагировали ни на какие экстероцептивные раздражения. Эти клетки распределялись по глубине расположения, как показано в табл. II.
Реакция спонтанно разряжающихся клеток на экстероцептивные раздражения обнаружена у 30 нейронов. Наибольшее число их (27) отвечало на соматосенсорные раздражения.
Из 27 нейронов, реагировавших на соматосенсорную стимуляцию, только на нее отвечало 19 клеток. Ответ выражался либо в начальном увеличении частоты разрядов (17 единиц), либо в их торможении (2 единицы).
Реакции этих нейронов наступали с разными латентными периодами. У 13 клеток ответ возникал с латентным периодом до 10 мсек. У 4 нейронов реакция на соматосенсорную стимуляцию наступала со скрытым периодом 15-20 мсек.
Спонтанно активные клетки, реагирующие на раздражение седалищного нерва или кожи, найдены преимущественно на глубине 550-2000 мк (см. табл. II).
Часть нейронов ядра клиновидного канатика реагировала не только на соматосенсорные стимулы, но также и на неспецифические для данного ядра раздражения. Таких полисенсорных нейронов в данном образовании было обнаружено 8, причем одни из них отвечали на соматосенсорные и звуковые раздражения (3 клетки), другие - на соматосенсорные, световые и звуковые воздействия (5 клеток).
Кроме того, у 3 нейронов, обследованных по неполной программе, найдены реакция только на звуковые (2 клетки) и только на световые (1 клетка) раздражения.
Из 8 единиц, обследованных по полной программе, у 8 были обнаружены изменения в характере активности в ответ на действие звуковых раздражений, у 5 - на световые стимулы и у 3 - на соматосенсорные раздражения.
Поливалентные нейроны найдены у животных, находящихся под хлоралозным наркозом на глубине до 1000 мк (1 клетка), на глубине 1050-1500 мк (5 клеток) и на глубине 1550-2000 мк (5 клеток) (см. табл. II).
Полисенсорные нейроны ядра клиновидного канатика реагируют на экстероцептивные раздражения учащением импульсации, торможением, а также двухфазными процессами, т. е. возрастанием ритма разрядов с последующим периодом молчания или наоборот. Иногда реакция была многофазной.
Из 28 эффектов, обнаруженных у поливалентных клеток, 5 были выражением усиления процесса возбуждения, 11 - торможения и 12 включали как тот, так и другой процесс.
Характер реакции этих нейронов на разные модальности раздражений представлен в табл. III.

Таблица III. Характер реакции нейронов ядра клиновидного канатика на раздражения разных модальностей
Анализ этих данных показывает, что начальные эффекты распределяются при действии стимулов разных модальностей следующим образом.
Латентные периоды ответов (усиления импульсации) полисенсорных нейронов на стимулы разных модальностей колебались в пределах до 10 мсек для соматосенсорных, в пределах 15-80 мсек для звуковых и 30-80 мсек для световых воздействий. Лишь у 1 нейрона скрытый период реакции на звуковые щелчки превысил 80 мсек. Более подробная расшифровка этих данных представлена в табл. IV. Из этой таблицы следует, что один нейрон реагировал начальным усилением импульсации и на специфические и на неспецифические (свет, звук) раздражения. У 3 нейронов реакции на все виды применяемых воздействий были тормозными. У остальных 4 единиц ответы были разнохарактерными.
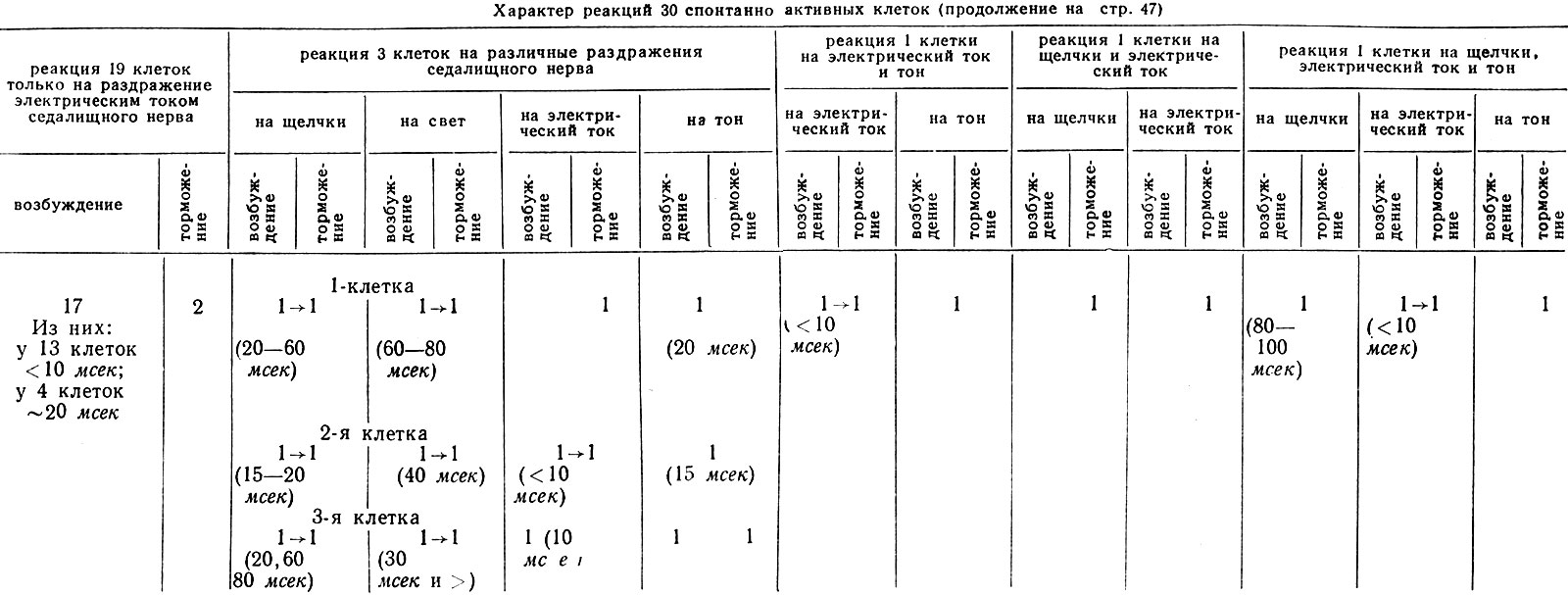
Таблица IV. Латентные периоды реакций нейронов ядра клиновидного канатика на раздражители разной модальности

Таблицы IV. Продолжение
Примечание: в скобках указана величина латентного периода реакции в мсек; стрелка - наличие двухфазных реакций.
Наибольшее количество полисенсорных нейронов (5) реагировало на раздражения 3 модальностей. Пример таких нейронов дается на рис. 10, где приводится одновременная запись разрядов 3 клеток, расположенных на глубине 1250 мк от поверхности клиновидного ядра кошки, находящейся под хлоралозным наркозом в сочетании с диплацином.
Как следует из рис. 10, а, через 80 мсек после включения щелчка появляется вспышка (40-50 в 1 сек) высокоамплитудных импульсов клетки № 1 (спайки 750 мкв), длящаяся 50 мсек, за которой следует период молчания (1 сек). Частота разрядов клетки № 2 (спайки 500 мкв) существенно не меняется. Действие света (см. рис. 10, б) не сопровождается существенными сдвигами в деятельности первой клетки, в то время как разряды второй клетки несколько учащаются. Раздражение седалищного нерва (см. рис. 10, в) в первой клетке вызывает вспышку импульсов в ритме до 70-90 в 1 сек с латентным периодом менее 10 мсек, за которой следует интервал молчания. Несколько урежаются в этот период и разряды второго нейрона. Включение тона (см. рис. 10, г) приводит к почти полному прекращению разрядов первого и второго нейрона. В то же время на фоне звуковой стимуляции происходит активирование третьего нейрона, частота разрядов которого возрастает почти вдвое.

Рис. 10. Разряды полисенсорных нейронов ядра клиновидного канатика кошки под хлоралозным наркозом в сочетании с диплацином. а - ответ на щелчки (60 дб); б - ответ на свет (300 лк, 0,1 мсек); в - ответ на раздражение седалищного нерва электрическим током (1 в, длительность импульса 0,1 мсек); г - ответ на тон (500 гц 50-70 дб). На рис. 10, а, б, в и следующих двух рисунках артефакт раздражения, плохо видимый на фотографиях, указан точкой над осциллограммами. На рис. 10, г отметка раздражения - опускание линии над осциллограммой. Глубина расположения клетки от поверхности ядра - 1250 мк (по Т. С. Наумовой и Н. А. Ласточкиной, 1966)
Нужно подчеркнуть, что характер реакции нейрона в большой мере определяется его функциональным состоянием. Ответная реакция клеток на неболевые стимулы зависит от того, например, применяется ли раздражение впервые или повторно.
Так, на рис. 11, где представлена осциллограмма клетки, расположенной на глубине 1600 мк (кошка под хлоралозным наркозом в сочетании с диплацином), видно, что каждый звуковой щелчок, следующий в ритме 4, а затем 2 в 1 сек, вызывает эффект, характер которого зависит от его порядкового номера в серии. Первые 3 щелчка сопровождаются возрастанием ритма разрядов с 25-30 до 70-80 в 1 сек. Реакция наступает с латентным периодом 20 мсек; 4-й, 5-й щелчки вызывают, наоборот, урежение спайков до 12-18 в 1 сек.

Рис. 11. Разряды полисенсорного нейрона ядра клиновидного канатика кошки под хлоралозным наркозом в сочетании с диплацином в ответ на действие серии щелчков (60 дб). Верхняя линия - отметка звуковых раздражений. Глубина расположения клетки от поверхности ядра 1600 мк (по Т. С. Наумовой и Н. А. Ласточкиной, 1966)
Особенно показательной явилась запись активности другого нейрона, расположенного на глубине 1250 мк, также полученная на кошке в состоянии хлоралозного наркоза в сочетании с диплацином (рис. 12). Действие первого щелчка (см. рис. 12, а) вызывает реакцию, состоящую лишь из эффекта возрастания ритма разрядов (более чем в 2 раза), наступающего с латентным периодом порядка 20 мсек. При втором применении этого раздражителя (тот же рисунок) длительность эффекта активации сокращается до 50 мсек, уступая место торможению, которое через 100 мсек опять сменяется некоторой активацией. Реакция наступает со скрытым периодом 50-60 мсек. Этот же нейрон реагирует на вспышки света также двухфазной реакцией, соотношение фаз в которой зависит от интервала между стимулами (см. рис. 12, б, 12, в). Скрытый период ответа также сокращается с 40 до 30 мсек. Раздражение седалищного нерва вызывает аналогичную реакцию, характер которой находится в зависимости от исходного состояния нейрона. Одна из этих реакций приведена на рис. 12, г. Она выражается в появлении вспышки разрядов (50 мсек), сопровождающейся длительным (более 500 мсек) периодом молчания клетки.

Рис. 12. Разряды полисенсорного нейрона ядра клиновидного канатика кошки под хлоралозным наркозом в сочетании с диплацином. а - ответ на щелчки (60 дб); б, в - свет (300 лк, 0,1 мсек); г - раздражение электрическим током (1 в, 0,1 мсек) седалищного нерва. Глубина расположения клетки от поверхности ядра 1250 мк (по Т. С. Наумовой и Н. А. Ласточкиной 1966)
Итак, 8% нейронов специфического ядра Бурдаха, этого первичного переключательного реле кожно-двигательного анализатора, проводящего тактильную и проприоцептивную чувствительность, реагируют не только на соответствующие соматосенсорные раздражения, но и на неспецифические для них звуковые и световые стимулы. Иными словами, это специфическое реле ствола мозга может проводить и неспецифическое возбуждение, и в нем могут осуществляться процессы конвергенции импульсов разных модальностей. Если сопоставить эти данные с данными, полученными при изучении активности нейронов ретикулярной формации, то получится что "неспецифических" элементов в специфических системах в 4-5 раз меньше, чем в ретикулярной формации, ибо, по данным Mancia и сотр., (1957), поливалентных клеток в ретикулярной формации насчитывается 37%, по данным Palestini и сотр. (1957), - 50%. Иными словами, различия в полисенсорных элементах специфических и неспецифических систем на уровне ствола мозга лишь количественные.
Встает вопрос о том, каковы структурные и функциональные особенности тех поливалентных нейронов ядра клиновидного канатика, которые реагируют не только на специфические, но и на неадекватные для этого ядра (световые и звуковые) раздражения. Известно, что основную массу клеток ядра клиновидного канатика составляют типичные чувствующие клетки, аксоны которых имеют коллатерали, обеспечивающие связь нейронов между собой, а также связь этих нейронов с соседними специфическими ядрами и ретикулярной формацией (Ramon у Cajal, 1909).
Кроме этих элементов, в состав ядра Бурдаха входят нейроны, которые по строению можно отнести к нейронам, образующим основную массу клеток ретикулярной формации (Lenhossek, 1895; В. М. Бехтерев, 1896, 1898; Ramon у Cajal, 1896; М. Е. Шайбель, А. Б. Шайбель, 1958; Г. П. Жукова, 1959; Т. А. Леонтович, 1959, и др.). Наибольшее количество нейронов ретикулярного типа находят в участках специфических ядер, прилегающих к ретикулярной формации (Т. А. Леонтович, 1959; Г. П. Жукова, 1959; Е. Г. Школьник-Яррос, 1959).
В наших опытах поливалентные нейроны ядра Бурдаха были обнаружены на глубине 1200-2000 мк от его поверхности, т. е. в участках ядра, близких к ретикулярной формации. Латентные периоды ответов таких нейронов на стимулы разных модальностей колебались в наших опытах в пределах от 2 до 10 мсек для соматосенсорных, в пределах 15-80 мсек для звуковых и 30-80 мсек для световых воздействий. Эти данные совпадают с результатами наблюдений Amassian и De Vito (1954), показавшими, что нейроны ретикулярной формации реагируют на экстероцептивные раздражения со скрытым периодом 12-80 мсек. Таким образом, по положению и по величине латентного периода полисенсорные нейроны сходны с нейронами ретикулярной формации. С другой стороны, величина скрытых периодов реакций поливалентных нейронов ядра Бурдаха на соматосенсорные стимулы (порядка 10 мсек) сближает их со специфическими элементами этого ядра. На основании всего того, что было изложено, трудно решить вопрос о структурной организации полисенсорных нейронов ядра клиновидного канатика. Для этого требуются дальнейшие исследования. Судя по латентному периоду реакций поливалентных нейронов, можно думать, что проведение неспецифического возбуждения к ядрам клиновидного канатика осуществляется по коллатералям волокон слухового и зрительного нервов, которые достигают этого ядра как непосредственно, так и опосредованно через ретикулярную и другие формации. Нет доказательств, что в этом процессе не участвуют вышележащие структуры, в частности кора головного мозга и особенно сенсомоторная зона коры, куда возбуждение может поступать по прямым связям анализаторов. Так, Ю. В. Бирючковым (1963) из лаборатории С. Б. Дзугаевой показаны прямые пути, в частности, между зрительным нервом и моторной корой, которые отходят от зрительного анализатора на уровне corona radiata и идут в составе других сенсорных систем на субкортикальном уровне.
Как показали наши эксперименты, характер ответа полисенсорных нейронов ядра Бурдаха на экстероцептивные раздражения менялся под влиянием текущей афферентации и, следовательно, определялся исходным функциональным состоянием клетки, а именно, в зависимости от порядкового номера раздражения и интервала между стимулами ответная реакция могла носить как характер возбуждения, так и торможения, равно как и признаки того и другого процесса. Интересно, что по мере повторения стимула обычно происходило сокращение латентного периода ответа. Если же ответная реакция носила двухфазный характер, то по мере повторения раздражений обнаруживался сдвиг в сторону процесса торможения.
Эти данные свидетельствуют о том, что и на уровне первичных переключательных реле анализаторов функциональный фактор определяет полисенсорность нейронов подобно тому, как он определяет ее и на кортикальном уровне (О. С. Виноградова и Д. Ф. Линдслей, 1963). Как показали исследования В. С. Русинова (1953, 1956, 1960), явление функциональной полисенсорности играет несомненную роль при установлении временных связей между взаимодействующими анализаторами. Очевидно, это касается как кортикальных, так и субкортикальных уровней анализаторов, что будет обсуждено в следующей главе.
Итак, нейроны ядра Бурдаха, ответственные за проведение тактильной и проприоцептивной чувствительности, реагируют не только на соответствующие соматосенсорные раздражения, но и на неспецифические для них звуковые и световые стимулы, т. е. на уровне первичного переключательного реле кожно-двигательного анализатора может осуществляться взаимодействие возбуждений, вызванных раздражениями разных модальностей. К сожалению, очень мало известно о процессах конвергенции импульсов на стволовых уровнях других специфических систем анализаторов, и этот вопрос требует дальнейшего тщательного изучения.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'