V. Моделирование некоторых принципов функциональной организации нервной системы
Разработка вопросов теории и практики применения искусственных самоорганизующихся систем, или "обучающихся автоматов", обусловила повышенный интерес специалистов точных наук и техники к проблемам обучения человека и животных, количественному анализу структуры и функций нервной системы.
Однако огромный материал, накопленный физиологией в. н. д., получен преимущественно в упрощенных частных условиях, и поэтому многие выводы этих исследований не могут быть непосредственно использованы для сопоставления с результатами, полученными на описанных выше моделях. Не существует, например, сопоставимых количественных критериев для оценки оптимальности поведенческих реакций организма, эффективности переработки информации в различных отделах нервной системы и некоторых других показателей, необходимых для оценки возможностей мозга как сложной управляющей системы в сравнении с автоматическими устройствами.
Теория обучения автоматов охватывает широкий круг вопросов, который имеет отношение по крайней мере к следующим этапам условнорефлекторного поведения: 1) активный поиск и различение каждого из условных и безусловных раздражителей; 2) определение статистической структуры безусловных раздражителей во времени и пространстве, а также по отношению к окружающим предметам или сигналам (условным раздражителям); 3) различение внутренних состояний отдельных элементов организма (автомата) и их частных задач, определяемых текущим моментом и прошлым опытом; 4) формирование и выполнение реакции, 5) контроль адекватности (эффективности) реакции, и т. д.
Понятно, что рассмотренные в предыдущих разделах модели нейронов и самоорганизующихся систем - логические сети, конечные автоматы - являются лишь весьма отдаленными аналогами структуры или функций нервной системы. Однако ценность моделей заключается не только в наиболее близкой имитации работы мозга, но и в возможности сравнить с тем, как он мог бы или как он не мог бы работать. Здесь вполне уместно привести слова И. М. Сеченова (1952): "Мысль о машинности мозга при каких бы то ни было условиях для всякого натуралиста клад. Он в свою жизнь видел столько разнообразных причудливых машин, начиная от простого винта до тех сложных механизмов, которые все более заменяют собой человека в деле физического труда, он столько вдумывался в эти механизмы, что если поставить перед таким натуралистом новую для него машину, закрыть от его глаз ее внутренности, показать лишь начало и конец ее деятельности, то он составит приблизительно верное понятие и об устройстве этой машины и об ее действии". Успехи современной электроники и развитие кибернетики позволяют создавать машины, заменяющих человека и в деле умственного труда, и нужно полагать, что эти устройства для изучения мозга окажутся еще более полезными, чем машины времен И. М. Сеченова.
Самое главное, что дали нам кибернетические представления о деятельности мозга, - это то, что важнейшая функция мозга - эффективное, целенаправленное управление и регуляция внутренних состояний в изменяющейся внешней среде - доступна для функционального моделирования, и что деятельность мозга подчинена тем же ограничениям, которые установлены теорией информации. Так, например, оптимальное приспособление живого организма, заключающееся в поддержании постоянства его основных параметров в допустимых пределах при наличии внешних изменений и вредных воздействий, гомологично с коррекцией шумов в канале связи.
По известной теореме следует, что для исправления ошибок путем некоторого числа выборов через корректирующий канал должно пройти соответствующее данному числу выборов количество информации. И мозг в этом случае действует так же, т. е. не может выполнить эти функции, пока не примет и не передаст необходимого количества информации (если не верить в чудеса). Данное обстоятельство, конечно, не следует понимать пессимистически, ибо таким же образом открытие закона сохранения энергии положило конец попыткам построить вечный двигатель, но оно же явилось началом бурного развития и успехов техники в новом направлении. Зная ограничения, которым подчиняется работа мозга, мы сможем глубже понять его механизмы и детали строения.
Вообще при изучении сложных систем, имеющих иерархию уровней ее организации или форм движения материи, следует всегда иметь в виду высшие функции системы или направления развития, которым подчиняются детали ее структуры. Поскольку можно предположить, что мозг является оптимальным регулятором, то целесообразно ознакомиться со структурами современных систем автоматического управления. Оставляя в стороне простые случаи, когда в системе имеется полная информация о состоянии объекта и возмущающих воздействиях, рассмотрим систему с неполной информацией и автоматическим поиском (Фельдбаум, 1964). Основой решения какой-либо задачи в неопределенной ситуации является метод активного поиска (Бернштейн, 1962; Гельфанд и соавт., 1962). Этот метод состоит в пробах различных путей и способов решения, без гарантии успеха. В простейшем случае (или когда имеется очень мало информации) используется метод слепого или случайного поиска (выбора), или известный в биологии метод "проб и ошибок" (Эшби, 1956). В последнее время большее внимание уделяется методам эвристического программирования, где используются психологические данные о действии человека по достижению какой-либо цели (Миллер, Галантер, Прибрам, 1959). Прежде всего выбирается (на основе прошлого опыта) зарекомендовавшая себя стратегия, которая проверяется в действии, и в случае неудачи вносится коррекция, выдвигается новая гипотеза о возможных путях решения задачи, и так далее до получения успеха (хотя и этот метод не дает полной гарантии успеха).
Рассмотрим схему автоматической оптимизации (рис. 93). Управляемый объект В находится под воздействием управляющего сигнала u от управляющего устройства А1 и случайной помехи z, медленно изменяющейся во времени. Выходная реакция должна повторять закон действия х′ задающего сигнала, причем во всех каналах связи имеются помехи, искажающие передаваемую информацию. Автоматический оптимизатор А2 воспринимает значения критерия оптимальности О и ограничений Hi, вычисленных устройством С на основании сравнения задающего и выходного сигналов. Затем, действуя через управляющее устройство на объект, автоматический оптимизатор изменяет параметры А1 в нужном направлении, в зависимости от реакции объекта, для повышения критерия оптимальности.
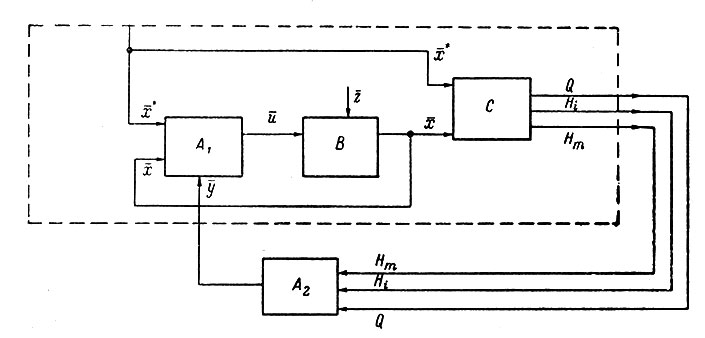
Рис. 93. Структурная схема оптимального управления с автоматическим поиском. A1- управляющее устройство; В - управляемый объект; С - вычислительное устройство; А2 - автоматический оптимизатор (прерывистой линией очерчен объект оптимизации). Объяснения в тексте
Вообще обычная система автоматического регулирования состоит из следующих основных элементов: входное и выходное устройства, переключательный блок (управления), цепи обратной связи и органы сравнения прямого и обратного сигнала, моделирующее устройство (в схемах управления по модели) и устройство памяти. Аналогичные структуры можно найти и в нервной системе (Поляков, 1964; Соколов, 1964б). На рис. 94 и 95 показаны схемы нейронной организации ориентировочного и условного рефлексов. Принципиальное отличие этой структуры заключается в пластичности ее элементов.
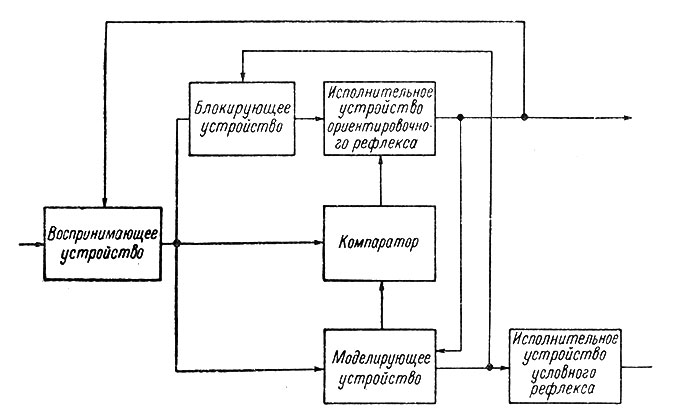
Рис. 94. Структурная схема ориентировочного и условного рефлексов. (Соколов, 1964б)
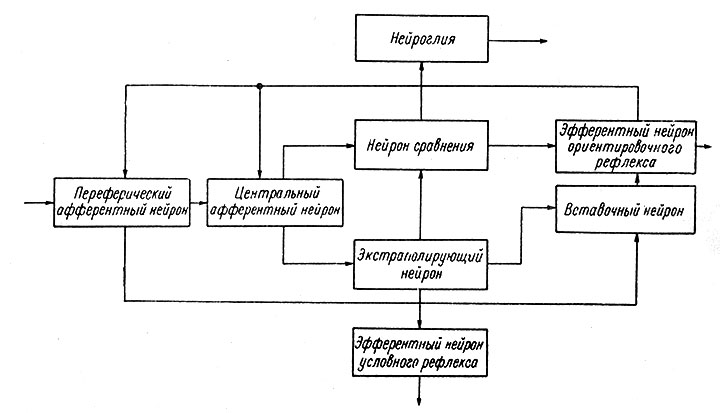
Рис. 95. Схема нейронной организации ориентировочного и условного рефлексов. (Соколов, 1964б)
По-видимому, в биологических системах управления и регулирования с целью наилучшей выживаемости в условиях изменяющихся в широких диапазонах влияний внешней среды и активной борьбы за существование в процессе эволюции вырабатывались очень гибкие, разнообразные механизмы перестройки не только в количественном (саморегуляция), но и в качественном (самоорганизация) аспектах. Поэтому можно предположить, что нервная система и ее элементы могут изменять свои свойства под влиянием биологически значимых (экологически адекватных) раздражителей внешней среды и внутренних состояний организма постольку, поскольку это необходимо для наилучшей выживаемости. В этом смысле быстрота реакций может выигрываться за счет точности и наоборот, а вычислительные функции могут "обмениваться" с логическими и т. д. Так, например, можно полагать, что нейрон или элемент такой информационной системы имеет весьма сложный алгоритм преобразования выходного сигнала как по самой функции входа, так и по ее интегралу и производной во времени и в пространстве для оптимального выполнения текущей задачи данного нейрона в составе целостной деятельности нервной системы. Этот, беспорядочный на первый взгляд, смешанный код оказывается наиболее эффективным при передаче и переработке биологически важной информации в прошлом, в настоящий момент и в будущем. Можно думать, что в нервной системе производится поэтапная выработка реакций с последовательными приближениями точности. Такой способ действия позволяет в общих чертах сформулировать реакции почти мгновенно и затем по мере необходимости их уточнять и детализировать, что дает возможность обеспечить гибкий компромисс между правильностью действия и постоянной готовностью к любым неожиданным раздражениям.
Можно предположить, что саморегуляция и самоуправление организма во внешней среде выполняются сложной иерархией управляющих систем на основе физических, логических, семантических и прагматических принципов функционирования. При этом последние являются высшими и управляют низшими, в соответствии с программой деятельности целого организма (Виннер, 1948; Эшби, 1956; Буллок, 1964; Поляков, 1964).
Казалось бы, можно построить теорию функционирования мозга, даже не зная внутреннего строения отдельных нейронов. Однако мы еще слишком мало знаем и об общих или основных принципах взаимодействия их. Отдельно исследовали электрические, химические и различные биофизические параметры отдельных нервных клеток и пытаются экспериментировать с группами клеток, контролируя правильность анализа тем, что объяснения работы и реакций ансамблей клеток не должны противоречить принципам, полученным при исследовании отдельных единиц.
Последние несколько десятилетий характеризуются представлениями о нервной системе как о некоторой весьма сложной телефонной сети с автоматическим коммутатором или обычной электронной вычислительной машине, в которой определенные виды сигналов или взаимодействий передаются по определенным каналам связи и поэтому могут быть точно локализованы в каждый данный момент. Однако в настоящее время все чаще подчеркивается существенное отличие структуры реальной нервной системы от обычной электрической сети с "сосредоточенными постоянными". Предполагается, что все основные функции мозга - способность к обмену веществ, энергией и информацией, память, переключения, классификация различения и тому подобное распределены по миллионам клеток, одновременно участвующих во множестве других системных процессов и составляющих статистическую систему переменных времени, места и состояния (мозаику возбуждений и задерживаний по И. П. Павлову). Такое представление отличается от структуры логической сети из дискретных элементов, и пока еще нет удовлетворительных моделей такого типа, но разработка отдельных элементов для этой цели продолжается.
Особенный интерес в нейрофизиологии и нейрокибернетике в настоящее время представляют различные модели и гипотезы о материальных носителях памяти, или следов возбуждений в нервной системе, на которых могут быть построены правдоподобные модели временной связи. Многочисленными исследователями разрабатывались модели памяти как циклической активности нервных импульсов в замкнутых нейронных контурах. Успехи математической модели взаимосвязи между внутриклеточными и внеклеточными значениями потенциалов действия (Ролл, 1961, и др.) дают основание надеяться, что эта модель может быть использована и при исследовании иных проблем нейрофизиологии, представляющих значительный интерес. Например, найденное различие между вялой реакцией на периферическую дендритную синаптическую активность и значительно более быстрой реакцией на синаптическую активность в соме (и стволе дендрита) позволяет провести обобщение ранее предложенных рефлекторных моделей типа "вход-выход". Это различие между синаптической активностью в дендритах и соме может оказаться также очень важным при рассмотрении проблемы нейронных взаимодействий в больших популяциях. Данный эффект должен быть учтен в коэффициентах, выражающих запаздывающую связь.
Другое направление в исследовании следов возбуждения в нервной системе относится к поискам физиологических механизмов адаптации, постетанической потенциации, роста отдельных элементов нейронов, проявляющихся в ходе "проторения" пути при выработке условного рефлекса.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'