VI. Вероятностные модели нейронной организации нервной системы
Сложность и многообразие изучаемых в биологии явлений, изменчивость полученных данных, вариаций параметров (когда, например, индивидуальные различия превышают видовые) привели ко все более широкому применению в биологических и медицинских исследованиях статистических методов оценки результатов на основе теории вероятности и информации. Однако значение статистических методов не ограничивается только методикой обработки наблюдений. Можно предположить, что в основе всей интегративной деятельности нервной системы большую роль играет статистический принцип (Walter, 1953). Это обусловлено прежде всего тем, что внешняя среда, в которой мы живем, в самом общем случае, является вероятностной средой, т. е. состоит не только из жестко детерминированных и повторяющихся (стереотипных) ситуаций, но и из случайных событий. Каждое из этих событий невозможно предсказать абсолютно точно, но их совокупности образуют более устойчивые явления и ситуации (системы событий). Поскольку организм непосредственно воспринимает информацию именно об отдельных событиях, то для оптимального "уравновешивания" и эффективной экстраполяции изменений внешней среды становится необходимым использование принципа статистической обработки сенсорной информации, своего рода фильтрации, выделения существенной информации и выявления стереотипов раздражителей.
Таким образом, в самом общем виде, мы можем представить себе, что в нервной системе на основе следов случайных раздражений организуется некоторая "статистически обработанная" модель внешнего мира (мозаика возбуждений и задерживаний), которая обеспечивает возможность подготовки будущих реакций на основе оценки вероятности событий по прошлому опыту.
Системность и динамические стереотипы условных реакций исследовались П. К. Анохиным, Э. А. Асратяном, П. С. Купаловым и многими другими, однако в этих работах степень детерминированности (стереотипности) систем раздражителей и реакций могла оцениваться только качественно. Лишь в последние годы в связи с применением теории информации в психологии (Бойко, 1964; Соколов, 1964б), появилась возможность количественной оценки разных степеней "упорядоченности стереотипа" в зависимости от вероятности появления отдельных сигналов или от энтропии (неопределенности) системы раздражителей в целом. В частности, было показано (Бирюков, 1963; Меницкий, 1964), что некоторые параметры двигательных рефлексов (скорость выработки и угасания, частота появления и стабильность) существенно зависят от величины логарифма вероятности подкрепления условного раздражителя безусловным (рис. 96). Показано также, что в Т-образном лабиринте животные (крысы) при соотношении вероятностей подкрепления 1:3 забегают только в одну сторону лабиринта. В наблюдениях за работой человека - оператора на пульте управления в аналогичной вероятностной внешней среде было показано, что человек оценивает не только среднюю вероятность ситуации в целом, но и вероятность отдельных событий в предъявляемой последовательности сигналов (опыты Б. Г. Будашевского). Эти исследования частично подкрепляют значение математических моделей "условной вероятности" и "условной достоверности" А. М. Аттли (1956), по также показывают недостаточность их определения (как отношения числа подкреплений к общему числу стимулов) без учета энтропии системы раздражителей.
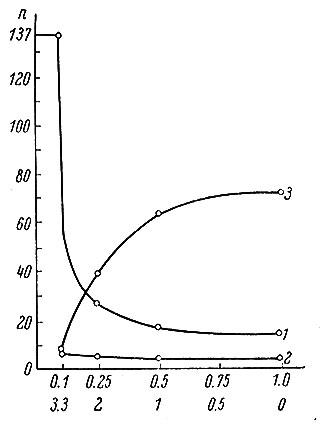
Рис. 96. График зависимости выработки двигательного условного рефлекса у рыб от вероятности электрического подкрепления. (Ловчиков и Меницкий, 1963). По оси абсцисс - уменьшение неопределенности подкрепления в битах (сверху) и увеличение вероятности подкрепления (снизу); по оси ординат - число предъявлений условного раздражителя. 1 - до первого проявления условного рефлекса; 2 - между отдельными проявлениями рефлекса в период его упрощения; 3 - кривая, обратная 1, показывающая логарифмическую зависимость скорости выработки рефлекса
При изучении внутренних физиологических механизмов и принципов организации центральной нервной системы мы также встречаемся со статистическими закономерностями. Прежде всего это выражается в нестабильности, флуктуации возбудимости отдельных нейронов (Вартанян и Василевский, 1964б) и других физиологических реакций (Меницкий, 1964).
С фактом огромной индивидуальной вариабильности организмов мы встречаемся во всех особенностях их макро- и микроструктуры. По каждой из морфологических особенностей может быть установлен свой вариационный ряд, или спектр градаций. Чем более высокодифференцированной является ткань, тем сильнее выражено своеобразие в строении каждого ее элемента. В нервной системе наблюдается исключительное многообразие нейронов по форме и размерам. К области индивидуальных вариаций должны быть отнесены и особенности протекания в каждой ткани разнообразных ферментативных процессов и участвующих в них ферментных систем, которые в своей основе едины для всех организмов (Поляков, 1964).
Таким образом, из всего вышесказанного можно сделать вывод, что флуктуация и нестабильность реакций являются существенной характеристикой нервной системы и ее элементов. Это дает основание рассматривать деятельность нервной системы на ее различных уровнях с точки зрения отношения полезного сигнала и помех (или шумов), которые до сих пор преднамеренно снижались условиями лабораторных опытов (использованием систем раздражителей с минимальной энтропией, т. е. стереотипов) или рассматривались как артефакт эксперимента.
На основании изложенных данных можно также обосновать положение о том, что понятие порога реакции в большинстве случаев следует рассматривать именно с вероятностной точки зрения. Увеличение интенсивности раздражителя повышает вероятность появления реакции, но при достаточно длительном испытании могут быть и реакции на подпороговые раздражители в обычном смысле слова.
Что касается реакций отдельных нейронов и их роли в реакции всей системы, то их можно рассматривать как некоторые случайные величины, зависящие от времени. Каждому нейрону соответствует определенная вероятность появления разряда или изменения частоты импульсации, причем выработка новых реакций всей системы перераспределяет как вероятности разрядов отдельных нейронов, так и вероятности включения их в разные оперативные совокупности (ансамбли по А. А. Ухтомскому) нейронов (Коган и Соколов, 1965).
В свою очередь, эти совокупности (нервные центры) находятся между собой в аналогичных отношениях и под влиянием потоков нервной импульсации образуют констелляции центров более высокого уровня интеграции. Осуществляется ли высшая интеграция в вероятностных ансамблях или в упорядоченных структурах обратной связи - этого мы пока не знаем. Во всяком случае на структуру вероятностной модели нервной системы не налагается существенных ограничений, кроме предположения, что связи нейронов могут быть совершенно случайными, а вероятность взаимодействия убывает с расстоянием. Эта динамическая организация нервных процессов поддерживается вероятностной сменой (перегруппировкой) элементов под влиянием обмена информацией (нервной импульсации).
Эти модельные представления, основанные также на анализе микроэлектродных исследований, позволяют высказать гипотезу о значении аритмичной фоновой активности нейронов как факторе поддержания тонуса центральной нервной системы в соответствии с принципом "оптимального взаимодействия центров" (Меницкий, 1964, 1965). Аналогичные соображения о необходимости некоторого среднего (оптимального) уровня афферентации высказывает и Р. Юнг (Jung, 1961). В вычислительных машинах шум поддерживается на минимальном уровне и в нормальных условиях не влияет на истинные интегративные функции. В отличие от этого фоновая активность нервной системы, имеет величину приблизительно того же порядка, что и сигналы, действующие на ее фоне. Она поддерживает средний уровень активности. Таким образом, фоновая активность, или нормальный нейронный "шум", несет положительную функцию.
Однако следует пояснить, что, хотя совершенно случайная структура является логически наиболее простой и обеспечивает выполнение любой схемы связи между ее элементами, она (случайная структура) не всегда будет самой экономной или целесообразной. В тех случаях, когда внешние воздействия или соответствующие реакции специализированы и стереотипны, более эффективной оказывается система с адекватно сформированной постоянной структурой (вероятность некоторых соединений выше, чем вероятность остальных). С развиваемой точки зрения специализация сложной системы - это ограничение степеней свободы (уменьшение энтропии состояний) данной системы. Специализация функций соответственно приводит к более жесткому ограничению (специализации) вероятностной структуры нервной системы (и обратно), причем интересные примеры подобной специализации сенсорных путей под влиянием ограничения разнообразия внешней среды (экологическими факторами) дает нам эволюционная и экологическая физиология нервной системы (Карамян, 1962; Бирюков, 1963; Поляков, 1964, 1965). Статистическая характеристика взаимодействия основных нервных процессов на различных уровнях интеграции позволяет установить связь классических понятий о возбуждении и торможении с данными современной нейрофизиологии центральной нервной системы (Клини, 1961).
Непосредственная приложимость понятия информации к изучению деятельности нервной системы следует из того, что возникающий уже в рецепторе нервный процесс и формирующийся на его основе психический процесс восприятия явным образом представляют собой различные частные формы информационных процессов (Веккер, 1964). Поэтому общие характеристики и особенности этих частных форм могут быть раскрыты лишь путем установления соответствующих различий в конкретных проявлениях общих закономерностей переработки, передачи и хранения информации. Нейрокибернетика вывела эти формы нервных процессов за пределы психофизиологии и нейрофизиологии, обобщила и систематизировала их, создав тем самым общую теорию информации и управления. Создание общего для живых организмов и других самоорганизующихся систем единого научного языка является важным достижением кибернетического синтеза, и основной предпосылкой для успешного моделирования структуры и деятельности функций центральной нервной системы человека и животных.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'