V. Особенности нейрональной организации отдельных анализаторов в связи с их эволюцией
Реакции отдельных нейронов в различных анализаторах во многом сходны, хотя они перерабатывают информацию различной модальности и имеют неодинаковую организацию периферических рецептивных полей и связей друг с другом. Сходство некоторых основных типов реагирования (on-, off-, on-off и др.), по всей вероятности, определяется происхождением всех отделов ц. н. с. из одного зачатка.
В процессе эволюции из первоначально разобщенных и разбросанных в эктодермальных тканях нервных клеток формируется диффузная, а затем и ганглиозная нервная система (Заварзин, 1941). На уровне позвоночных на базе диффузной и ганглиозной нервной системы складывается ц. н. с. и ее разнообразные отделы. В ходе эволюции ц. н. с. прогрессивно увеличивались не только ее масса и количество отдельных нейронов, но и главным образом развивались и усложнялись связи между отдельными нейронами. В свою очередь усложнение связей сопровождалось специализацией и дифференциацией нейронов.
Однако в ходе специализации и дифференциации в системах нейронов, а также, возможно, и в отдельных нейронах произошло как бы наслоение эволюционно новых механизмов регуляции их деятельности на эволюционно более старые формы управления.
В нервной системе развитие интегральных механизмов шло от диффузных, неспецифических форм к локальным, специализированным типам реагирования (Орбели, 1933, 1941; Бирюков, 1947, 1948, 1960, и др.). Эти две стадии формирования нервных механизмов имеют, вероятно, и определенную нейрофизиологическую основу или определенные типы функциональной организации нервных элементов, начиная от рецепторов и кончая центральными образованиями. Отражением постепенного эволюционного формирования интегральных механизмов в центральной нервной системе является наличие двух нейрональных систем передачи и анализа чувствительной информации: неспецифической малоспециализированной и малодифференцированной и специфической, в высшей степени дифференцированной и специализированной. А. И. Карамян (1964) считает особенно плодотворным анализ функций нервной системы с точки зрения баланса специфических и неспецифических механизмов.
Г. И. Поляков (1964) также рассматривает прогрессивную дифференцировку анализаторных систем как переход от диффузных взаимосвязей различных уровней к более организованному характеру переключений. В настоящее время можно утверждать, что этот процесс затронул практически все анализаторы.
В кожном анализаторе экстралемнисковая система передачи является эволюционно более ранней, доказательством чего, по мнению Херрика и Бишопа (1958), является малый диаметр нервных волокон в этой системе. Различие двух систем кожного анализатора проявляется также и в неодинаковой степени морфологической дифференциации отдельных нервных клеток (Бехтерев, 1905; Жукова, 1964; Поляков, 1964, и др.).
В зрительном анализаторе древнюю систему передачи, по-видимому, составляет палочковый аппарат сетчатки, не имеющий спектральной специфичности, а филогенетически новую систему представляет колбочковый аппарат, обладающий избирательной спектральной чувствительностью. Анализ этих систем по диаметру афферентных волокон затруднен, поскольку первичные афферентные нейроны находятся в сетчатке, а зрительный нерв является в сущности центральным трактом. Выделение в зрительном анализаторе специфической и неспецифической систем в настоящее время представляется затруднительным.
Еще большие трудности вызывает попытка выделить элементы двойственной организации сенсорной системы слухового анализатора. Девис (1961) предлагает рассматривать улитку и ее афферентные пути как филогенетически новый орган, а отолитовый прибор и полукружные каналы как эволюционно более древнюю систему восприятия и анализа низкочастотных механических вибраций.
В области внутреннего анализатора также выявляется две системы передачи: по системе блуждающего нерва и по чревному нерву. Чревный нерв содержит толстые быстро проводящие волокна, а блуждающий - тонкие волокна. Соответственно различаются и их центральные проекции: у блуждающего нерва они диффузны, у чревного - локальны (Дуринян, 1966).
Эволюционные преобразования афферентных систем проявляются своеобразным образом в экологических изменениях их строения и функций. Так, например, в кожном анализаторе наиболее плотная сеть рецептивных окончаний и волокон обнаружена в тех участках кожного покрова и слизистых, которые наиболее часто взаимодействуют с окружающей средой и при этом извлекают наиболее существенную информацию о ней. Соответственно и проекции этих афферентных систем в центральную нервную систему имеют ряд особенностей. Многими авторами (Penfield a. Rassmussen, 1950; Woolsey, 1958, и мн. др.) показано, что размеры корковых зон проекций отдельных участков кожной поверхности неодинаковы. Зоны, связанные с экологически значимыми рецептивными полями (например, у животных область морды, дистальных отрезков лап; у человека - кожная поверхность губ, большого и указательного пальцев, и т. д.), больше тех участков коры, куда поступает афферентная импульсация от менее значимых зон.
Анализ рецептивных полей у отдельных корковых нейронов в различных зонах проекции кожного анализатора также выявил ряд примечательных особенностей. Так, у кошек нейроны с локальными рецептивными полями одной модальности встречаются главным образом в области представительства кожных рецепторов морды и дистальных отрезков лап (особенно передних). Размеры рецептивных полей нейронов, связанных с рецепторами кожной поверхности туловища, в среднем во много раз превосходят величину рецептивных полей только что указанной группы нейронов (Mountcastle, 1957; Anderson, 1962; Darian-Smith, 1964). Только во второй соматосенсорной зоне у кошек значительная часть нейронов (примерно 1/3) имеет распространенные рецептивные поля. Такого типа клетки в первой соматосенсорной зоне встречаются в меньшем количестве. В целом соматосенсорная кора кошек является высокодифференцированным образованием, содержащим нейроны преимущественно с локальными рецептивными полями. Обнаружены некоторые особенности организации рецептивных полей у корковых нейронов кроликов. В зоне SI нейроны с локальными рецептивными полями встречаются практически только в области представительства рецепторов головы и шеи. В области представительства кожной поверхности туловища и лап такого рода нейроны встречаются реже, причем их рецептивные поля больше, чем у кошек в этой же области соматосенсорной зоны коры. Если у кошек у части клеток размеры рецептивных полей нейронов, связанных с рецепторами дистальных отрезков передних лап, могут быть равными нескольким десяткам квадратных миллиметров, то у кроликов, как правило, рецептивные поля охватывают либо кожную поверхность целого диетального сегмента конечности (3-4 см2), либо всю поверхность конечности. В зоне представительства туловища и лап сравнительно часто встречаются нейроны с распространенными рецептивными полями и конвергенцией афферентных влияний различной модальности (рис. 80). На рис. 81 показаны количественные соотношения между нейронами разных типов в различных участках соматосенсорной области кроликов. В целом зона SI у кроликов по организации связей отдельных нейронов весьма близка ко второй соматосенсорной области кошек.
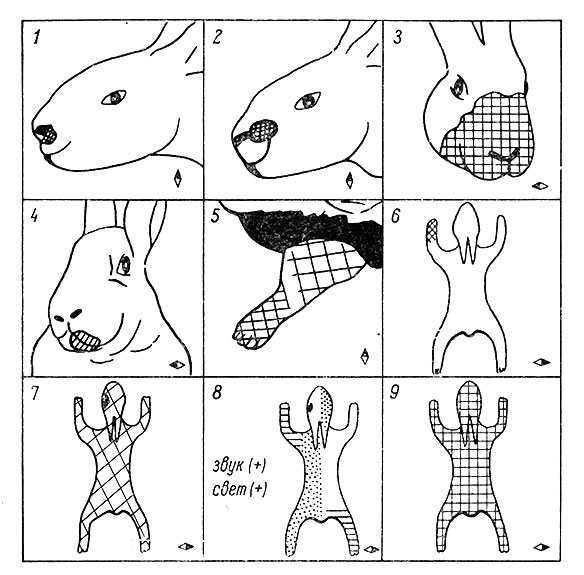
Рис. 80. Примеры организации рецептивных полей в различных участках коркового представительства кожного анализатора у кроликов. (Ориг.). 1, 2, 3, 4, 7 и 9 - возможные типы нейронов в области представительства кожной поверхности головы: 1,2,4 - нейроны с локальными рецептивными полями, 3,9 - нейроны с распространенными рецептивными нолями на контра- и ипсилатеральной поверхности кожи, 7 - нейрон переходного типа с коротколатентным локальным и длиннолатентным распространенным рецептивным полем. 5, 6 и 8 - возможные типы нейронов в области представительства туловища и конечностей: 5, 6 - нейроны с локальным рецептивным полем с тормозной примыкающей каймой (5) и без нее (6), 8 - нейрон с полимодальным рецептивным полем, реагирующим на звук, свет, болевые раздражения (точки) и давление (горизонтальная штриховка). Затушеванная половинка ромба - указатель стороны отведения в коре больших полушарий. Для 1, 2 и 5 отведение - с контрлатерального полушария мозга
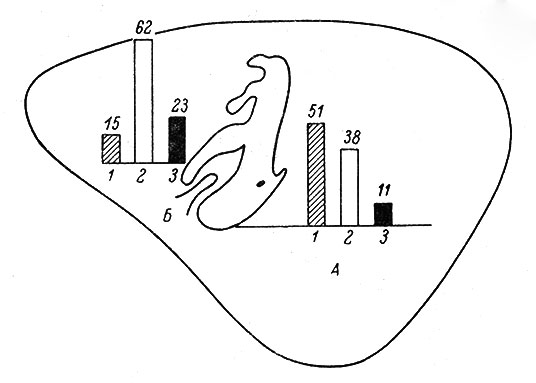
Рис. 81. Количественные соотношения между различными типами нейронов в отдельных участках первой соматосенсорной зоны коры кроликов. (Ориг.). А - в области представительства кожной поверхности головы, Б - туловища и конечностей. 1 - нейроны с локальными рецептивными полями; 2 - нейроны с распространенными рецептивными полями; 3 - нейроны, у которых рецептивное поле не выявлено. Цифры над столбиками - процент от общего числа обследованных клеток в данном участке коры
Преобладание нейронов с распространенными рецептивными полями в коре у кроликов, по-видимому, объясняется не только экологическими особенностями организации соматосенсорной зоны но и тем, что у кроликов в коре отсутствуют ассоциативные зоны (Gazard a. Buser, 1963), где у других млекопитающих (кошка, обезьяна и др.) сосредоточена основная масса нейронов с широкой конвергенцией разнообразных афферентных путей (Baser а. Imbert, 1961, и др.). Вероятно, у кроликов неспецифические ассоциативные нейроны рассеяны и частично включены в функциональную структуру соматосенсорной зоны. Примечательно наличие в коре кроликов своеобразной группы клеток, которые можно отнести к переходной форме от нейронов неспецифического типа к мономодальным нейронам локального типа. У этих нейронов обширное рецептивное поле сочетается с наличием в нем зоны, раздражение которой сопровождается в несколько раз большим возбуждением.
Эти особенности организации центральных нейрональных систем кожного анализатора кролика в какой-то мере объясняют диффузный (общеповеденческий) характер условных двигательных реакций при нанесении безусловного болевого раздражителя на конечности (Бирюков, 1947, 1948; Климова, 1948). В то же время у этих животных описаны условные специализированные локальные реакции в виде дерганья зубами кольца (Малиновский, 1957).
В ходе эволюции имеет место прогрессивная дифференциация и специализация афферентных и двигательных систем передней конечности, которая достигает наивысшего совершенства у человека.
Как проявляется действие экологических факторов на таких этапах эволюции нейрональных механизмов, когда еще не выражена топическая специализация в отдельных образованиях центральной нервной системы? Для ответа на этот вопрос Н. Н. Василевский (1966) изучил особенности связей у нейронов кортикоидной пластинки больших полушарий голубей. Как известно, в коре птиц еще отсутствует топическое представительство отдельных анализаторов (Kappers, Huber a. Crosby, 1936). Было показано, что корковые нейроны имеют преимущественно полимодальные связи со зрительным, кожным, проприоцептивным и звуковым анализаторами. Выявилась своеобразная частость обнаружения этих связей у изученных групп нейронов корковой пластинки (рис. 82). Такое распределение частостей может найти объяснение в различной экологической значимости отдельных видов раздражений. Есть все основания считать, что интеграция в корковой пластинке зрительных, соматических и кожных раздражений является важнейшей в координации полета, разнообразных движений и других рефлекторных влияний. Ранее проведенные опыты Т. М. Загорулько (1955), В. И. Багрянского (1959) и А. Б. Когана (1956, 1964) также показали большое влияние зрительных раздражений на биоэлектрическую активность больших полушарий голубей и легкость образования на них условно-рефлекторных связей. Эти авторы подчеркивают важную экологическую роль света для голубей и значительно меньшее значение звуковых раздражений.
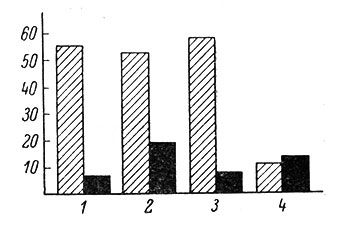
Рис. 82. Распределение частостей выявления вызванной активности у отдельных корковых нейронов голубя на зрительные (2), тактильные (2), проприоцептивные (3) и слуховые (4) раздражения. (Василевский, 1966). По вертикали - процент нейронов (от общего числа исследованных клеток), реагирующих на раздражение соответствующей модальности. Заштрихованные столбики - нейроны с возбудительными реакциями; черные столбики - с тормозными
Другие исследования, проведенные на нейронах гиппокампа рептилий - образования. Генетически близкого к кортикоидной пластинке голубей (Kappers, Huber a. Crosby, 1936) также показали экологическую специфику связей у отдельных нейронов. К. Г. Гусельникова и В. И. Гусельников (1963) и Л. Г. Воронин и сотрудники (1964) отметили большую эффективность соматических и зрительных раздражений по сравнению со звуковыми.
Итак, на уровне таких сравнительно мало дифференцированных образований головного мозга, как кора птиц и гиппокамп рептилий, полифункциональные связи у отдельных нейронов обнаруживают определенную направленность по отношению к экологически значимым раздражениям.
Таким образом, нейрональные организации афферентных систем проявляют в одних случаях высокую специализацию и дифференциацию связей, в других на фоне полифункциональных взаимоотношений наблюдается лишь определенная направленность связей. Эти особенности организации нейрональных систем определяются эволюционными механизмами и экологической специализацией вида.
У высших животных в нервной системе сохраняются эволюционно более древние нейрональные организации. Наслоение новых нейрональных афферентных и эфферентных систем сопровождается существенными изменениями функций нервной системы в целом. Между древними и новыми структурами устанавливаются теснейшие функциональные связи, причем новые системы функционально преобладают над эволюционно более старыми (Орбели, 1933, 1941; Карамян, 1964).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'