III. Слуховой анализатор
Импульсация, возникающая в рецепторах кортиева органа, передается через спиральный ганглий, слуховые ядра ствола и промежуточного мозга к височным слуховым полям коры больших полушарий.
Кацуки и сотрудниками (Katsuki et al., 1959) было показано, что по мере приближения к коре рецептивное поле (здесь - полоса пороговых звуковых частот, на которые реагирует нейрон) в целом сужается, однако наиболее узкую частотную полосу имеют нейроны медиального коленчатого тела. У кортикальных нейронов эта полоса несколько шире. Данное обстоятельство связывается с тем, что на корковом уровне процессы синтеза и интеграции приобретают ведущую роль. Наблюдаемое сужение полосы частот, по мнению Кацуки, связано с включением специальных тормозных механизмов, посредством которых вытормаживаются как более низкие, так и более высокие частоты, примыкающие к воспринимаемому диапазону. Можно полагать, что вытормаживание в значительной мере обусловлено кортикофугальными механизмами, а также взаимодействием нейронов на каждом уровне. Автор показал, что тормозящее влияние "соседних" диапазонов частот усиливается по мере приближения к коре.
Определение так называемых характеристических кривых (пороговых величин для разных частот) у отдельных нейронов различных уровней слухового анализатора выявило ряд интересных особенностей. Так, оказалось, что первичные нейроны имеют несимметричные и симметричные кривые. При несимметричной кривой порог изменяется постепенно при переходе от низких к более высоким тонам, на высоких тонах падает более круто, затем резко обрывается при переходе к предельно высоким частотам (рис. 79). Симметричная кривая выходит более равномерно в области высоких и низких частот. Кацуки (Katsuki, 1961) полагает, что эти два типа пороговых кривых определяются активацией различных групп волосковых клеток (наружных и внутренних).
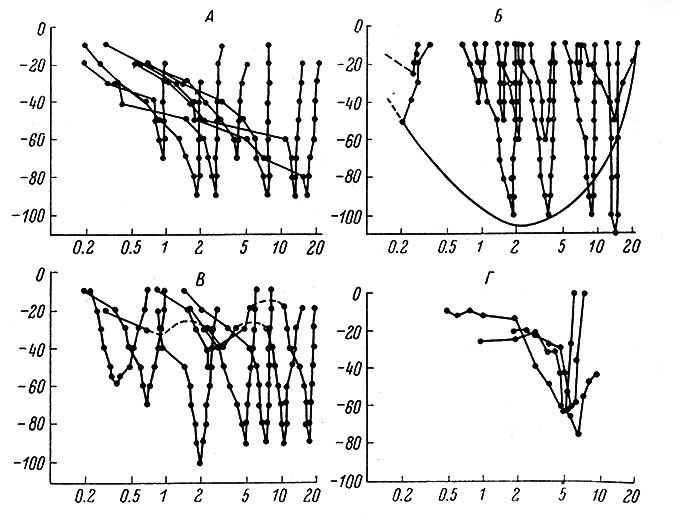
Рис. 79. Рецептивные зоны одиночных нейронов слуховой системы (Katsuki, 1961). А - нейроны, составляющие кохлеарный нерв; Б - нижнее двухолмие; В - трапециевидное тело; Г - медиальное коленчатое тело. По оси абсцисс - в логарифмическом масштабе частота, в кгц; по оси ординат - интенсивность звука, в дб
У каждого нейрона в слуховом нерве можно найти свою характерную форму реакции в зависимости от частоты и силы раздражения (Rupert et al., 1963). При этом по характеру разрядов нейроны делятся на две группы: быстро достигающие постоянного уровня вызванной активности при той или иной высоте раздражающего звука и медленно настраивающиеся.
При нарастании силы звука у периферических нейронов частота импульсации увеличивается и следует в основном S-образной зависимости. Такие силовые отношения отмечаются до субкортикального уровня; у корковых нейронов силовая зависимость представляется более сложной.
На уровне нижних бугров четверохолмия впервые появляются ответы типа on-off и частота их обнаружения увеличивается по мере приближения к коре.
В коре наиболее подробно анализ нейрональных реакций был проведен Кацуки и сотрудниками (Katsuki et al., 1959, 1961), Рупером и сотрудниками (Rupert et al., 1963), Нельсоном и сотрудниками (Nelson et al., 1963), Эвансом и Вайтфильд (Evans a. Whitfield, 1964). Было показано, что нейроны, воспринимающие высокие частоты, преимущественно располагаются в передней части g. ectosylvius; нейроны с рецептивными полями в среднем диапазоне частот - в средней ее части, а клетки с низкочастотным рецептивным полем - в задней трети этой извилины.
Эванс и Вайтфильд (Evans a. Whitfield, 1964) обнаружили специальную группу клеток, реагирующую только на резкую смену частоты звука. Выявлены также нейроны, возбуждающиеся как на звук, так и на движение предметов в поле зрения.
Важной особенностью нейронов слухового пути является снижение уровня фоновой активности по мере приближения к коре, в которой уже большинство нейронов не имеет фоновой активности. Кацуки (1961) было обнаружено, что нейроны в поверхностных слоях коры имеют быструю адаптацию и реагируют преимущественно на короткие звуковые раздражения; противоположный характер реакций проявляют более глубоко расположенные нейроны. Хюбел и сотрудники (Hubel et al., 1959) описали в слуховой коре так называемые клетки "внимания", реакция которых угасает при повторных применениях раздражителя.
Исследование второй слуховой корковой зоны (АII) обнаружило реакции нейронов, чрезвычайно напоминающие ответы ретикулярных нейронов ствола и таламуса: более высокий порог, отсутствие частотной специфичности рецептивного поля, более медленная адаптация, высокий уровень фоновой активности, наличие разрядов последействия раздражителя. Можно думать, что эти нейроны представляют собой неспецифические элементы коркового конца слухового анализатора - аналогично таким же элементам в других отделах коры во вторых проекционных зонах (см. ниже).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'