II. Зрительный анализатор
От фоторецепторов, через ганглиозную систему сетчатки по зрительному тракту афферентные импульсы достигают таламических релейных и неспецифических ядер, откуда сигналы поступают в кору больших полушарий к первой и второй зрительным проекционным зонам. Нейрональная организация зрительного анализатора представляется особенно многообразной и сложной. Разнообразие зрительной информации о яркости, свете, контрасте, движении, форме, цвете и других обеспечивается многочисленными нейрональными системами, тесно взаимодействующими друг с другом. Стабилизация зрительного образа и правильная его проекция в окружающую среду достигаются взаимодействием зрительной афферентации с вестибулярным, кожным двигательным и слуховым анализаторами.
В настоящее время детально описаны реакции отдельных нейронов на всех уровнях зрительной системы (Jung, 1961; Baumgartner, 1961, и мн. др.). Наибольшее многообразие реакций отмечается на корковом уровне. Так, было отмечено, что на подкорковом уровне у нейронов преобладают сравнительно однообразные концентрические рецептивные поля, что, возможно, связано с участием этих нейрональных комплексов преимущественно в релейных механизмах. В корковом конце зрительного анализатора, помимо рецепторных полей, сходных с полями подкоркового уровня, большинство клеток имеет несимметричные, распространенные поля, подчас занимающие всю поверхность сетчатки. Особенно сложные и широко охватывающие различные области сетчатки рецептивные поля имеют нейроны, относящиеся к системе восприятия движения (Jung, 1964). Баумгартнер и сотрудники (Baumgartner et al., 1964) полагают, что нейроны - "детекторы направления" являются исключительной принадлежностью коры больших полушарий. Нейроны, избирательно реагирующие при определенном направлении движения предмета в зрительном поле, значительно реже встречаются на уровне ганглиозного аппарата сетчатки (Barlow, Hill a. Levick, 1964). По мнению ряда авторов (Kozak et al., 1965), в функциональной структуре зрительной коры нейрональные системы контраста, движения и направления преобладают над системой восприятия яркости.
Типы нейронов в зрительной коре и других образованиях отражают стабильные отношения и внутренние свойства нейрональных систем, а также особенности их связей с ретинальными рецепторами. Надо особо подчеркнуть, что ответы нейронов зрительного анализатора могут быть поняты лишь при анализе самой зрительной информации. Все многообразие реакций наблюдается только при адекватном раздражении рецепторов (Jung, 1961). При всех других видах стимуляции, например при электрическом раздражении таламуса, ответы корковых нейронов совершенно не похожи на реакции при естественной засветке сетчатки (Akimoto u. Creutzfeldt, 1958; Creutzfeldt и. Akimoto, 1958). Вследствие сложной организации рецепторных полей классификация отдельных клеток возможна только после применения всего комплекса адекватных раздражителей (точечная и диффузная засветка, контраст, движение объекта и др.).
При монокулярной засветке сетчатки в корковом конце зрительного анализатора выделено 5 основных типов нейрональных реакций (рис. 78).
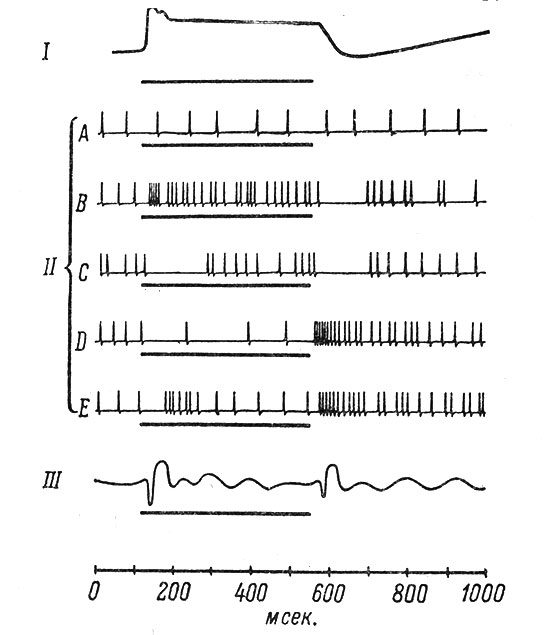
Рис. 78. Пять типов реакций нейронов зрительной коры на раздражение светом и темнотой и их связь с возбуждением рецепторов и ЭЭГ. (Jung et al., 1961). I - потенциал рецептора, регистрируемый внутриклеточным микроэлектродом, введенным в слой колбочек сетчатки; II - схематическое изображение разрядов различных типов нейронов; III - корковые потенциалы с эффектами включения и выключения, регистрируемые макроэлектродом с поверхности коры (ЭЭГ)
Нейроны типа А, составляющие 49% всех нейронов зрительной коры, не реагируют на однократные диффузные засветки сетчатки или на выключение света. Однако эти нейроны отвечают на вестибулярные раздражения и электрическую стимуляцию таламуса (Baumgartner u. Jung, 1955; Jung и. Baumgartner, 1955; Виноградова и Линдслей, 1963, и др.). Часть нейронов этого типа может быть активирована ритмическими повторными засветками сетчатки, движущимися объектами в зрительном поле (Hubel, 1959), применением светового контраста (Baumgartner u. Hakas, 1962) ИЛИ электрическим раздражением зрительного нерва (Jung, 1961). Однако даже при супрамаксимальном раздражении зрительного нерва у 28% кортикальных нейронов не наблюдается ответной реакции (Crutzner et al., 1958). При комбинации электрического раздражения с засветкой сетчатки удается понизить процент неактивирующихся нейронов до 16%.
Юнг (Jung, 1958) полагает, что благодаря стабильной группе нейронов А поддерживается необходимый "средний" уровень возбуждения в коре и что эти нейроны могут относиться к специальной резервной системе, функции которой точно еще не выявлены.
Нейрон типа В (on-элемент, элемент включения) реагирует возбуждением на включение света и торможением - на его выключение. Противоположный характер реакции обнаруживает нейрон типа D (off-элемент, элемент выключения). Нейрон типа Е (on-off-элемент, элемент включения-выключения) возбуждается как при включении, так и при выключении света. Противоположен ему нейрон типа С, реагирующий торможением на включение и выключение раздражителя.
Примечателен тот факт, что тип ответной реакции зависит от места приложения раздражителя на сетчатке. Так, было отмечено, что если при стимуляции одного глаза у нейрона выявляется С-тип реагирования, то при освещении другого глаза может быть В-тип и т. д. (Grusser u. Crusser-Cornehls, 1960). Таким образом, тип реакции не определяется внутренними свойствами нейронов, а связан с особенностями их афферентных связей.
Частота импульсации нейронов типа В обнаруживает логарифмическую зависимость от интенсивности светового раздражения. Частота разряда D-нейронов не имеет такой однозначной зависимости от интенсивности предшествующего светового раздражения, однако торможение этих нейронов, вызванное светом, проявляет логарифмическую зависимость от силы светового раздражения (Baumgartner, 1961).
Изложенные факты позволяют считать, что нейроны, реагирующие по on- и off-типу, находятся в реципрокных отношениях. Можно полагать, что возбуждение одних нейронов через систему вставочных нейронов приводит к торможению других.
Некоторые авторы (Jung, 1964) полагают, что нейроны on-типа передают информацию "светлее", а off-типа - "темнее". Однако, по мнению Баумгартнера (Baumgartner, 1961), это заключение справедливо только в том случае, если оба типа нейронов связаны с одним и тем же участком рецептивного поля сетчатки.
Кодирование световых контрастных отношений уже осуществляется на уровне нейронного аппарата сетчатки. Баумгартнер изучил особенности нейронов сетчатки и показал, что при точечном световом раздражении центра рецептивного поля активируются on-нейроны; раздражение периферии рецептивного поля тормозит их активность. Напротив, нейроны off-типа тормозятся при освещении центра рецептивного поля и активируются при световом раздражении его периферии. При проекции на сетчатку белых и черных полос наблюдаются характерные изменения активности on- и off-нейронов. Максимальная активность on-нейрона наблюдается в том случае, когда центр рецептивного поля находится у края белой полосы на границе ее с черной. Если центр рецептивного поля находится на середине белой полосы, то активность нейрона несколько снижается. Если на центр рецептивного поля проецируется темная полоса, то активность нейрона тормозится.
Нейроны off-типа из системы контраста максимально активируются при засвете сетчатки, если центры их рецептивных полей находятся в центре темной полосы.
Специальную систему составляют нейроны восприятия и анализа направления движения (М-система), выделенные из нейронов группы А. Необходимо подчеркнуть, что эта система нейронов не связана с анализом световых контрастов как таковых (Jung, 1964). Большинство нейронов М-системы проявляет активность при движении глаза в каком-то одном направлении. По данным Грюссер и сотрудников (Grusser et al., 1963), в реакции на движущийся объект истинным раздражителем является изменение его положения в рецептивном поле (а не контраст или контур объекта), поскольку было обнаружено, что у всех элементов перемещение раздражителя к центру рецептивного поля всегда более эффективно, чем к его периферии.
Изучение спектральной чувствительности рецептивных полей у отдельных нейронов зрительной коры показало, что существуют две группы клеток, одна из которых не проявляет избирательной чувствительности к какой-либо части спектра. Другая группа клеток реагирует на определенную длину световой волны. Отдельные подгруппы этих клеток проявляют наивысшую чувствительность в оранжево-красной (600-640 ммк), зеленой (520-540 ммк) и голубой (460 ммк) части видимого спектра (Motokawa et al., 1962). Третья часть клеток реагировала на свет противоположных участков спектра; реже встречались клетки, отвечающие на свет соседних участков. При этом с одного участка спектра может наблюдаться реакция по on-, с другого - по off-типу, и наоборот. Здесь нейроны находятся также как бы в реципрокных функциональных отношениях (de Valois a. Jones, 1961, и др.).
Своеобразными нейрональными системами зрительной коры являются так называемые клетки "внимания", реакция которых проявляется только на "новизну", на первые предъявления раздражителя (Виноградова и Линдслей, 1963).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'