I. Анализаторы кожной и проприоцептивной чувствительности
В кожном и двигательном анализаторах, как и во всякой афферентной системе, можно выделить два пути передачи афферентной импульсации: специфический ("лемнисковый") и неспецифический ("экстралемнисковый"). В лемнисковой системе имеет место высокая специализация и дифференциация функциональных свойств отдельных нейронов. Нейроны имеют рецептивные поля сравнительно небольших размеров и определенной модальности. Расположение рецептивных полей постоянно. Афферентная импульсация передается с коротким латентным периодом (8-30 мсек.). Такая характеристика функциональных свойств отдельных нейронов справедлива для лемнисковой системы по крайней мере высокоразвитых млекопитающих (кошка, обезьяна, человек). У низших млекопитающих (грызуны) организация лемнисковой системы, в частности проекционных корковых зон, несколько иная (см. ниже стр. 196).
В системе медиального и тригеминального лемнисков кожного анализатора проводится информация главным образом от поверхностной и глубокой тактильной чувствительности. Корковая зона лемнисковой системы была подробно изучена Маунткастлом (Mountcastle, 1957). В полях 2 и 3 выявлены преимущественно нейроны, имеющие рецептивные поля в коже; в поле 1 преобладают клетки, связанные с рецепторами глубоких тканей (Mountcastle a. Powell, 1959a, 1959b, и др.).
Нейроны поверхностной тактильной чувствительности реагируют на легкие касания к коже или к волосам по on- или on-off-типу (рис. 73) и проявляют быструю адаптацию. Нейроны, связанные с рецепторами глубокой тактильной чувствительности, возбуждаются при давлении на кожу; реакция отмечается в течение всего периода действия раздражителя и соответствующие нейроны медленно адаптируются (рис. 74).Сравнительно редко встречаются нейроны, реагирующие на вибрацию кожи и подкожной клетчатки, а также на температурные раздражения (Mountcastle, 1957, 1959, 1961а, 1961b; Gordon a. Paine, 1960; Gordon, 1964; Gordon a. Jukes, 1964a, 1964b; Darian-Smith, 1964, и др.).
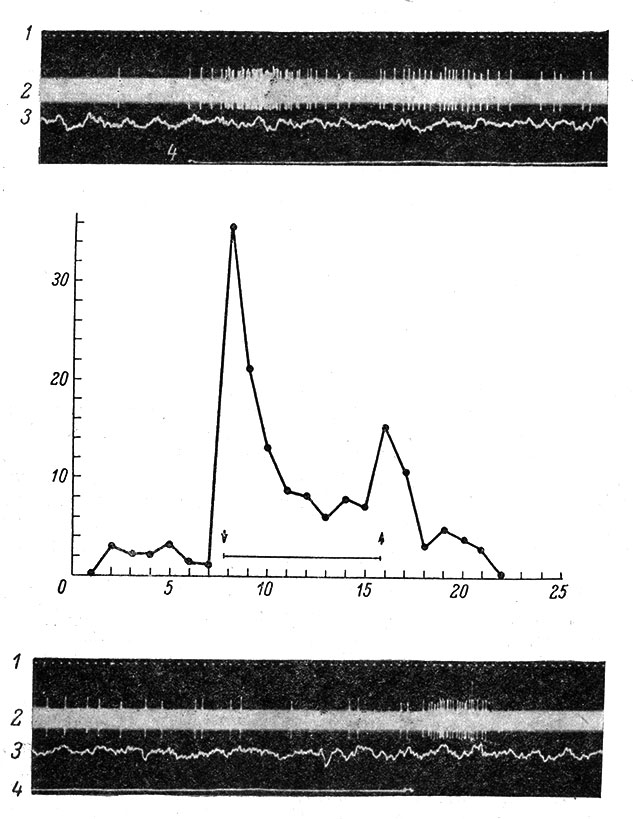
Рис. 73. Реакция быстроадаптирующегося нейрона в соматосенсорной коре кролика на длительное легкое прикосновение кисточкой к центру рецептивного поля. (Ориг.). Отмечается значительное учащение импульсации в момент прикосновения (верхняя осциллограмма) и обратного движения кисточки (нижняя осциллограмма). Кисточка приводилась в движение с помощью специального пневмомеханического устройства . 1 - отметка времени 1/20 сек.; 2 - внеклеточная электронейронограмма (ЭНГ); 3 - поверхностная ЭКоГ вблизи места погружения микроэлектрода; 4 - отметка раздражения. Калибровки: для ЭКоГ - 100 мкв, для ЭНГ - 5 мв. По оси абсцисс - время, в сек.; по оси ординат - частота импульсации, в имп./сек. Стрелки - начало и конец тактильного раздражения
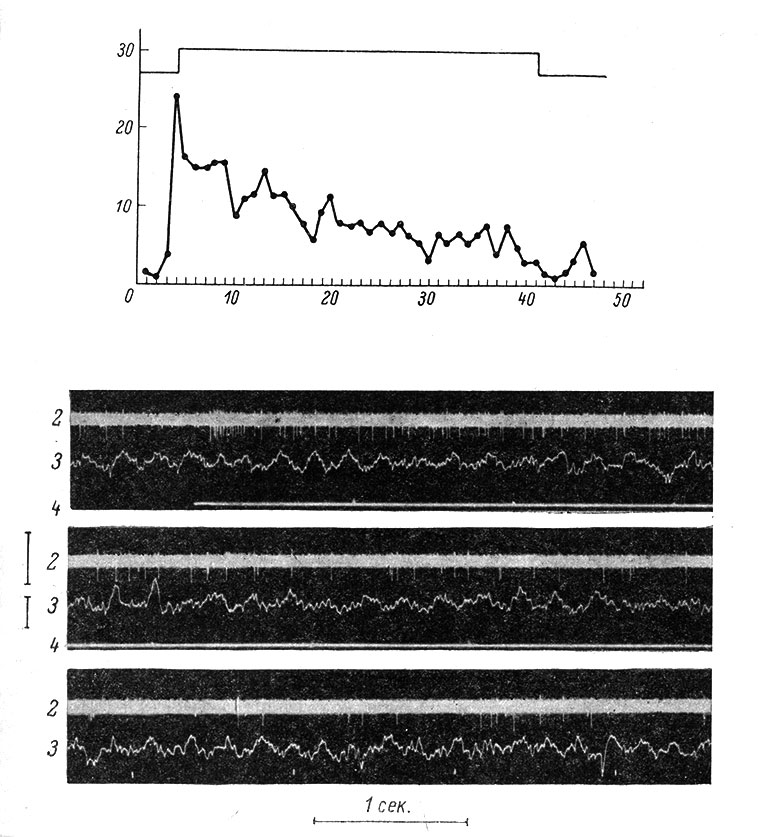
Рис. 74. Реакция медленно адаптирующегося нейрона в соматосенсорной области коры больших полушарий кролика на длительное постоянное давление кисточкой в центре рецептивного поля. (Ориг.). Осциллограммы, соответствующие моментам: включения раздражителя (верхняя), действия раздражителя, перед его выключением (средняя), после выключения раздражителя (нижняя). Остальные обозначения те же, что на рис. 73
Волокна и нейроны проприоцептивной чувствительности, относящиеся к медиальному лемниску, также характеризуются высокой степенью специализации рецептивного поля, - как в отношении модальности, так и в отношении его расположения. Выявлено два типа нейронов: нейроны "положения" и нейроны "движения". У первого типа нейронов реакция градуальна и зависит от угла, под которым конечность согнута в суставе (рис. 75). Большинство нейронов "положения" возбуждается либо только во время сгибания, либо только во время разгибания; однако у части нейронов реакция проявляется как при согнутой, так и при разогнутой конечности (Mountcastle a. Powell, 1959а, 1959b). У второго типа - нейронов "движения" - реакция наблюдается только в момент перемещения конечности. Реакция нейронов "положения" слабо адаптируется. У нейронов "движения", напротив, отмечается быстрая адаптация.
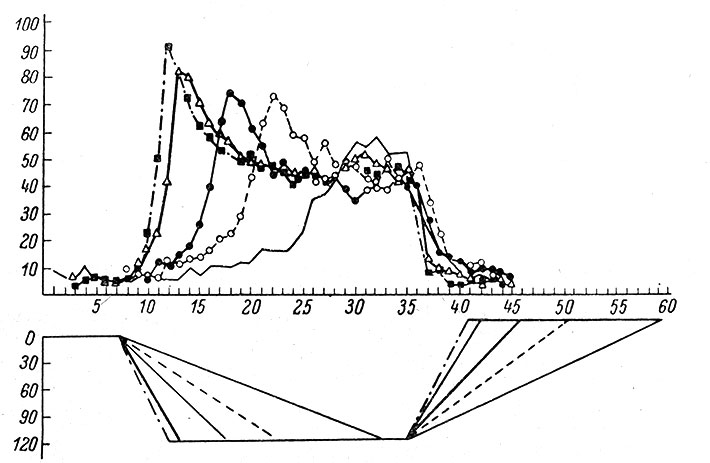
Рис. 75. Реакция нейрона 'положения' в зависимости от скорости сгибания и разгибания конечности в лучезапястном суставе. (Mountcastle, Poggio a. Werner, 1963). По оси абсцисс - время, в сек.; по оси ординат - вверху частота, в имп./сек., внизу - угловые градусы. Нижний график отражает 5 разных скоростей сгибания (разгибания) до одного и того же конечного положения в суставе; на верхнем графике видна соответствующая каждой скорости частота импульсации одного и того же нейрона
В экстралемнисковой системе (тонкие афферентные волокна, серое вещество спинного мозга, спинно-таламический путь, задняя группа ядер таламуса, вторая соматосенсорная область, отчасти и область SI) организация рецептивных полей у отдельных нейронов имеет более сложный характер. Рецептивные поля, как правило, обширны, нередко полимодальны, и вызванная реакция проявляется со значительным латентным периодом (30-100 и более мсек.). Ответы наблюдаются на термические, болевые и повреждающие раздражители. Следует отметить, что в данной системе особенности организация рецептивных полей связаны не только с особенностями первичных афферентных волокон, но и с тем, что уже на уровне нейронов второго порядка отмечается широкая конвергенция первичных ипси- и контрлатеральных афферентов (Kolmodin a. Skoglund, 1960; Уолл, 1961, и др.).
Корковая зона проекции экстралемнисковой системы (SII) имеет некоторые особенности по сравнению с SI. Значительная часть нейронов зоны SII реагирует на тактильные раздражения кожи и давление и имеет такие же локальные рецептивные поля, как и у нейронов вентро-базального ядерного комплекса таламуса. По всей вероятности, эти нейроны синаптически связаны друг с другом. 16% нейронов зоны SII отвечает на звуковые раздражения, причем у половины из них наблюдается вызванная реакция и на другие модальности раздражителей (Garreras a. Anderson, 1963). Около 10% нейронов в зоне SII отвечает на сильные раздражения глубоких тканей (мышц, связок, фасций, суставов; Anderson, 1962). В отличие от зоны SI, в зоне SII не обнаружены клетки, возбуждающиеся при легких движениях в суставах (Carreras a. Anderson, 1963). Как уже отмечалось выше, такого типа клетки встречаются только в системе медиального лемниска. Есть основания полагать, что нейроны в зоне SII, связанные с рецепторами глубоких тканей, получают информацию от высокопороговых волокон, иннервирующих суставы и, как известно, выделенных в самостоятельную группу Экклсом и Лундбергом (Eccles a. Lundberg, 1959).
Около 20% нейронов (Carreras a. Anderson, 1963), а по данным Н. Н. Василевского около 43%, составляют клетки с распространенными рецептивными полями, нередко с конвергенцией афферентных влияний различной модальности (рис. 76). Возможно, что характер рецептивного поля у них определяется проекцией волокон не только из задней группы ядер таламуса, но и из некоторых диэнцефальных неспецифических структур (Rose a. Woolsey, 1958; Anderson, 1962).
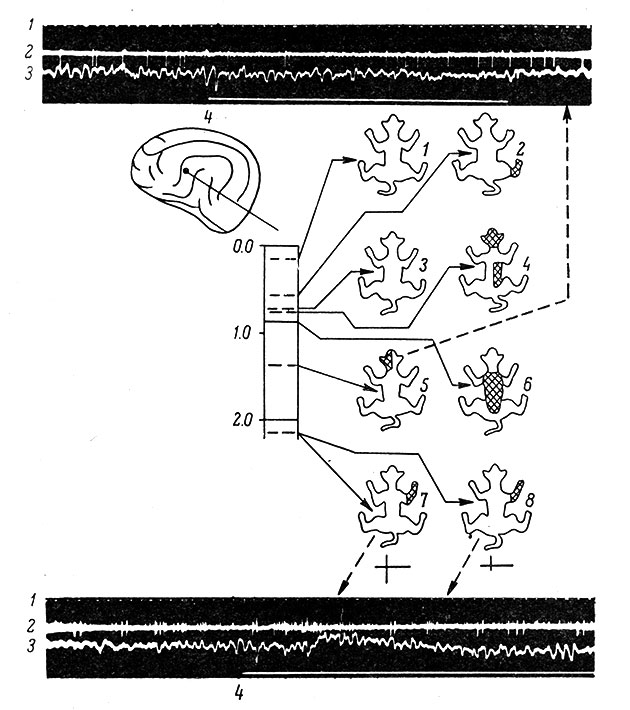
Рис. 76. Организация рецептивных полей у отдельных нейронов во второй соматосенсорной области коры одной кошки. (Ориг.). Слева на схеме показан пункт погружения микроэлектрода в g. ectosylv. ant. и уровни расположения нейронов в толще коры на данном треке электрода (микроэлектрод вводился под косым углом к поверхности коры). Цифры па схеме - глубина погружения микроэлектрода, в мм. Справа двумя вертикальными рядами представлены схематические изображения поверхности тела кошки, на которых заштрихованы рецептивные поля кожи, тактильное раздражение которых вызывает реакцию соответствующего нейрона (стрелка от схемы). 1 - нейрон, у которого не выявлено рецептивное поле; 2, 7, 8 - нейроны с локальными рецептивными полями; 3 - нейрон, реагирующий только на звук; 4,5 - нейроны с распространенными рецептивными полями; 6 - нейрон с рецептивным полем на ипсилатеральной поверхности морды. Внизу и вверху приведены осциллограммы, показывающие особенности реакций на тактильное раздражение. На нижней осциллограмме вызванная реакция появляется с коротким латентным периодом и представляется в виде быстро убывающей высокочастотной серии импульсов, а на верхней осциллограмме наблюдается слабая реакция, проявляющаяся лишь в некотором уплотнении межимпульсных интервалов. На осциллограммах: 1 - отметка времени (1/20 сек.), 2 - импульсная активность одиночного нейрона; 3 - ЭКоГ; 4 - отметка раздражения
Нейроны с распространенными рецептивными полями подразделяются на несколько подтипов: а) механорецептивные, б) реагирующие на сильные повреждающие воздействия и в) мультимодальные (тактильные и звуковые, тактильные и повреждающие или звуковые и повреждающие).
Выше уже отмечалось, что импульсация от проприорецепторов достигает центральных образований головного мозга по двум системам: через медиальный лемниск и через систему спинальных восходящих трактов, главным образом идущих к мозжечку (кунеоцеребеллярный пучок, дорсальный и вентральный спиноцеребеллярные тракты). Поскольку в последнее время было показано, что мозжечок имеет обширные связи с новой корой (Фанарджян, 1964), то обе системы передачи в конечном итоге конвергируют к корковым полям двигательного анализатора.
Особенности нейрональной организации восходящих спиноцеребеллярных путей были подробно изучены Оскарсоном (Oscarsson, 1956, 1957), Оскарсоном, Розеном и Удденбергом (Oscarsson, Rosen a. Uddenberg, 1963), Оскарсоном и Розеном (Oscarsson a. Rosen, 1963), Холмквистом и сотрудниками (Holmquist et al., 1956, 1963а, 1963b) и др.
Двигательная область занимает несколько цитоархитектонических полей в коре больших полушарий, например у кошек и обезьян, главным образом поля 4 и 6. Как указывалось выше, значительное число нейронов "движения" и "положения" встречается также и в области постцентральной извилины: в полях 2 и 3 и особенно в поле 1. Очевидно, следует рассматривать корковое представительство двигательного анализатора более обширным, чем это ранее считали многие авторы (Гамбарян, 1962; Саркисов, 1964, и др.).
В двигательной коре обнаружены полисенсорные нейроны, отвечающие на любые соматические, звуковые и зрительные раздражения, а также моносенсорные клетки, реагирующие с обширных или локальных рецептивных полей (рис. 77) (Buser a., Imbert, 1961). Показано, что моносенсорные нейроны с локальными рецептивными полями являются проекциями лемнисковых путей и относятся к специфической проприоцептивной системе. Нейроны с распространенными рецептивными полями и полисенсорные клетки, по-видимому, получают импульсацию как по восходящим неспецифическим, так и по интракортикальным путям.
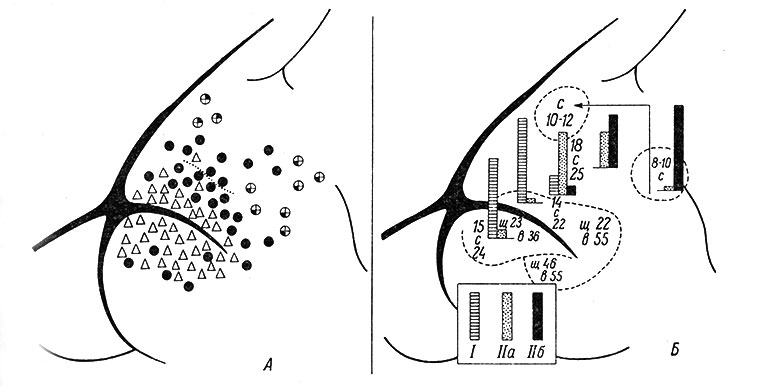
Рис. 77. Схема распределения в двигательной коре нейронов, относящихся к различным группам в соответствии с их 'чувствительностью' к периферическим раздражителям. (Buser a. Imbert, 1961). А - общая топография; каждая 'точка' представляет по крайней мере пять зарегистрированных клеток; треугольники - полисенсорные элементы (группа I); черные кружки - соматические элементы с распространенными рецептивными полями (группа IIа); перекрещенные кружки - соматотопические нейроны (группа IIб), реагирующие на стимуляцию контрлатеральной задней или передней конечности; прерывистая линия - теоретическая задняя граница поля 4. Б - гистограмма распределения клеток групп I, IIа и IIб, зарегистрированных в передней сигмовидной извилине и позади крестовидной борозды. Цифры показывают приближенные значения пределов латентных периодов для соматических раздражителей (с), вспышек света (в) и звуковых щелчков (щ). Глубокий хлоралозовый наркоз
Брукс, Рудомин и Слеймен (Brooks, Rudomin a. Slayman, 1961а, 1961b) выделили нейроны с фиксированными и лабильными перемещающимися рецептивными полями. При этом в моторной коре преобладают нейроны с фиксированными рецептивными полями (60% спереди и 80% сзади от s. cruciatus). Две трети нейронов с фиксированными рецептивными полями имели локальные рецептивные поля, располагающиеся на контрлатеральной стороне тела. Размеры рецептивного поля были наименьшими в дистальном отрезке конечности и прогрессивно увеличивались по мере приближения к локтевому или коленному суставам (с 2-4 мм2 до 8 см2). Форма рецептивного поля чаще была овальная или округлая, вытянутая вдоль конечности.
Нейроны с фиксированными распространенными рецептивными полями в 3/4 случаев имели поля на обеих сторонах тела, у 1/2 нейронов поля были специфическими по модальности. У 2/3 нейронов рецептивные поля были симметричными.
Только у 10% нейронов с лабильными рецептивными полями отмечается локальность их расположения, в остальных случаях имеются распространенные рецептивные поля.
Интересно отметить, что в моторной коре кошек только 10-20% нейронов связаны с рецепторами, реагирующими при движении конечностей. Основная масса клеток возбуждается на кожные, зрительные, слуховые, и другие модальности стимулов (Brooks, Rudomin a. Slayman, 1961a). Однако Альб-Фессар (Albe-Fessard, 1964) описывает в моторной коре обезьян нейроны, преимущественно имеющие связи с рецепторами глубоких тканей. Можно полагать, что в различных участках моторной зоны имеются неодинаковые количественные соотношения между отдельными типами нейронов.
Своеобразные связи имеют нейроны двигательной коры с вестибулярным анализатором. Так, Корнхубер и Ашов (Kornhuber a. Aschoff, 1964) наблюдали ответы на вестибулярные раздражения у 53% нейронов. Большинство нейронов активировалось при катодическом раздражении ипсилатерального лабиринта и тормозилось при анодической его поляризации. Следует подчеркнуть, что специфическую вестибулярную афферентацию получают только нейроны, связанные с рецепторами глубокой чувствительности; причем эти связи выявлены у 2/3 нейронов "положения". У нейронов с кожными рецептивными полями наблюдались вариабиальные и менее определенные реакции на вестибулярные раздражения.
Таким образом, моторная кора представляет собой своеобразную функциональную структуру, у которой, с одной стороны, имеются обширные полисенсорные афферентные связи, и она является местом конвергенции разнообразных модальностей раздражителей, напоминая ассоциативные области коры; с другой стороны, она имеет локальные афферентные проекции к двигательным ядрам ствола спинного мозга. Кстати, данная точка зрения уже высказывалась в литературе (Батуев, 1964, 1966).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'