IV. Вызванная активность центральных нейронов
Если ф. а. является в основном отражением функционального состояния мозга, то показателем распространения в ц. н. с. эффектов действующего раздражения является вызванная активность.
В различных сенсорных и других функциональных системах мозга типы вызванной активности разнообразны. Наиболее простым часто встречающимся типом вызванной активности для всех систем является ответ на включение раздражителя (on-эффект, эффект включения), который проявляется с некоторым латентным периодом в виде одного импульса или высокочастотной пачки пиковых потенциалов, содержащей от 1 до 28 пиков. Количество импульсов в пачке зависит от силы применяемых раздражений, локализации раздражений в рецептивном поле, а также от особенностей той или иной функциональной системы. Так, ответы клетки Реншоу могут содержать до 28 (чаще 8-10) пиков (J. С. Есcles, R. M. Eccles, Iggo a. Lundberg, 1961), в коре число пиков в пачке не превышает 3-5 (Amassian, 1961).
После завершения on-эффекта импульсная активность нейрона частично или полностью возвращается к исходному уровню, несмотря на продолжающееся действие раздражителя, а иногда даже тормозится.
При увеличении силы раздражения наряду с увеличением числа пиков в пачке и их плотности укорачивается латентный период (рис. 66). При этом с усилением раздражения вначале укорочение латентного периода идет быстро, а затем более медленно. Укорочение латентного периода наиболее отчетливо наблюдается в полисинаптических цепях проведения, что, возможно, связано с однонаправленной синхронизацией активности нейронов на каждом уровне проведения.
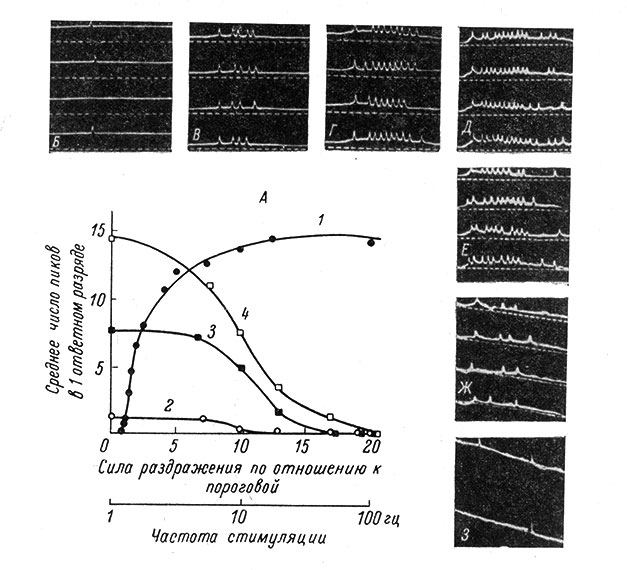
Рис. 66. Зависимость ответной реакции промежуточного нейрона от интенсивности и частоты раздражения афферентного мышечного нерва. (Вартанян и Василевский, 1963а). А: 1 - зависимость между интенсивностью раздражения (частота 1 гц) мышечного нерва и средним числом пиковых потенциалов в ответном разряде промежуточного нейрона; 2-4 - влияние частоты стимуляции на ответный разряд (2 - при пороговой силе раздражения, 3 - при силе раздражения в 2 раза выше пороговой, 4 - при силе раздражения в 20 раз выше пороговой). Б-З - фрагменты осциллограмм ответных разрядов того же нейрона при пороговой силе раздражения (Б); при силах раздражения в 2.5 (Б), в 3 (Г) и в 20 раз (Д) превышающих пороговую; Е-З - то же, что и Д, но при частоте 5 гц (Е), 20 гц (Ж) и 50 гц (З). Запись внеклеточная. Момент нанесения раздражения совпадает с началом развертки. Отметка времени 1 мсек
В отдельных случаях при увеличении силы раздражения отмечается удлинение латентного периода, что объясняется включением высокопороговых тормозных систем (Kennedy a. Towe, 1958).
При увеличении частоты раздражения число импульсов в пачке во всех случаях уменьшается по кривой, близкой к экспоненциальной (рис. 66). Однако при этом общее число импульсов, генерируемых нейроном в единицу времени, нарастает и в целом наблюдается как бы эффект облегчения. Такое поведение проявляется как у нейронов спинного мозга (Василевский, 1962; Вартанян и Василевский, 1963а, 1964а, 1964б), так и у нейронов коры (Mountcastle a. Powell, 1959a; рис. 67).
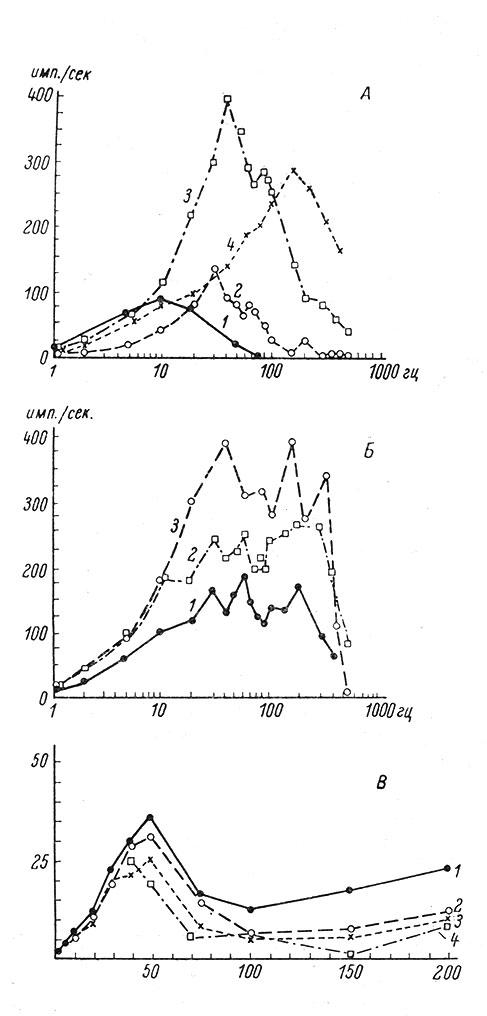
Рис. 67. Изменение суммарной импульсной активности нейронов спинного мозга (А, Б - по Н. Н. Василевскому, 1964) и коры (В - по Mountcastle a. Powell, 1959b) при увеличении частоты электрических раздражений афферентных волокон. По оси абсцисс - частота раздражения, в имп./сек. (на А, Б - в логарифмическом, на В - в линейном масштабе); по оси ординат - частота импульсной активности в 1 сек., в гц. А - суммарная импульсная активность вставочных клеток переднего рога, Б - клеток Реншоу, В - коры. 1-4 - примеры отдельных нейронов (первая, вторая, третья и четвертая секунды при каждой частоте раздражения)
В сенсорных системах только что описанные закономерности вызванной активности (особенно на повторные раздражения) представляются гораздо сложнее. Многими исследователями было показано, что величина вызванной активности, а также ее латентный период в значительной мере определяются локализацией места приложения раздражителя внутри рецептивного поля данного нейрона. Так, для кожного анализатора наибольший ответ отмечается при раздражении центра рецептивного поля; по мере приближения его к краям ответная реакция уменьшается, а латентный период увеличивается (рис. 68). В отдельных случаях отмечено, что раздражение участков кожи, примыкающих к рецептивному полю, дает тормозный или облегчающий эффект (Gordon a. Seed, 1961; Gordon a. Jukes, 1962, и др.). Этот эффект, по нашим наблюдениям, отмечается главным образом у нейронов поверхностной тактильной чувствительности.
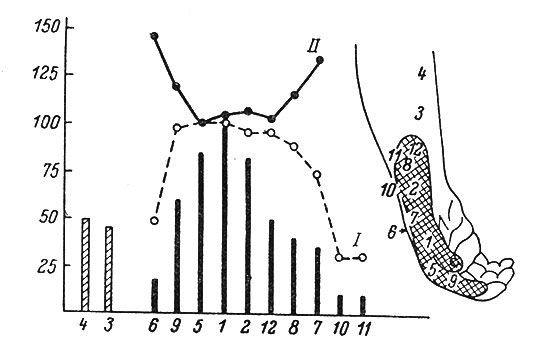
Рис. 68. Величина вызванной реакции нейрона постцентральной извилины макака при раздражении различных участков (1-12) рецептивного поля кожи. (Mountcastle a. Powell, 1959b). По горизонтали - номера, соответствующие одноименным участкам рецептивного поля; по вертикали - параметры реакции в данном участке (в % по отношению к участку 1). I - вероятность появления реакции; II - средний латентный период группы реакций. Раздражения, наносимые в 3 и 4, находящихся вне рецептивного поля, определяемого при электрическом раздражении, как полагают авторы, как-то активируют нервные волокна, иннервирующие поле
Реакция на включение является наиболее общим типом реагирования, поскольку она отражает наиболее простые функциональные взаимоотношения между центром и периферией. Этот вид реакции встречается во всех сенсорных системах.
Выделены два вида реагирования по on-типу: с быстрой и с медленной адаптацией (например, для кожных рецепторов). on-эффект у нейронов неспецифического типа (см. главу 8) может переходить в длительное последействие (рис. 69).
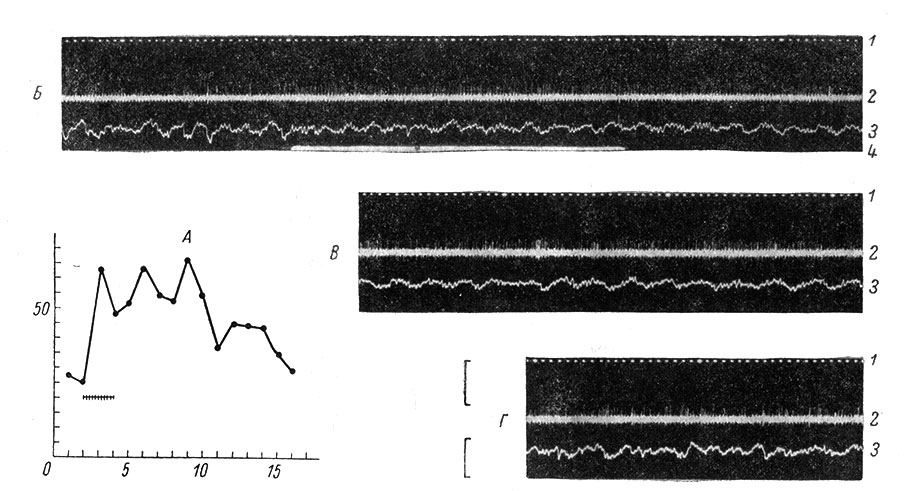
Рис. 69. Особенности реакции коркового нейрона с распространенным рецептивным полем неспецифического типа на кратковременное ритмическое тактильное раздражение. (Ориг.). На графике А по оси абсцисс - время, в сек.; по оси ординат - частота, в имп./сек.; внизу - отметка раздражения. Видна длительная следовая реакция нейрона; Б - осциллограмма, соответствующая 1-5 сек. графика; В - 8-10 сек.; Г - 15-16 сек.; 1 - отметка времени 1/20 сек.; 2 - ЭНГ; 3 - ЭКоГ; 4 - отметка тактильного раздражения; калибровка для ЭНГ - 5 мв, для ЭКоГ - 100 мкв
Наряду с нейронами, реагирующими по типу on, описаны off-нейроны ("элементы выключения"), которые реагируют на выключение раздражителя, а на включение его затормаживаются. Известны также нейроны on-off-типа, активируемые как включением, так и выключением раздражителя. Описаны и другие типы реакций, одни из которых являются стабильными формами ответа нейрона, а другие меняют свой тип реагирования в зависимости от условий раздражения. Последнее имеет место, например, в коре при замене диффузного раздражения сетчатки светом на мелькающий контрастный свет или движущийся раздражитель (Hubel, 1959; Hubel a. Wiesel, 1959; Чораян, 1963б, и др.).
В коре и подкорковых неспецифических ядрах описаны нейроны, активность которых проявляется только на "новизну" раздражителя (Hubel, 1959; Виноградова и Линдслей, 1963; Jasper, 1964, и др.). Такого же типа клетки в кортикоидной пластинке голубей наблюдал Н. Н. Василевский (1966).
Вызванная активность, как и фоновая, является флуктуирующей, нестабильной реакцией нейрона. В связи с этим ряд авторов выдвинул так называемый принцип неопределенности синаптического проведения (O'Leary, 1962, и др.), объясняющий относительную стационарность вызванной реакции нейрона.
Изменчивый флуктуирующий характер вызванной активности нейрона нередко проявляется среди значительной фоновой его импульсации ("шумов"). Если при этом имеется еще и большой латентный период, то при обычной визуальной оценке подчас бывает трудно решить, есть ли вообще вызванная активность. Герштейн и Киянг (Gerstein a. Kiang, 1960) отмечали, что при тщательном анализе активности корковых нейронов число клеток "без ответа" значительно уменьшается. В настоящее время имеется тенденция ко все более широкому использованию статистических методов для углубленного количественного анализа активности отдельных единиц.
Широкое распространение получило составление гистограмм межимпульсных интервалов. Герштейн и Киянг предложили два типа гистограмм: общую гистограмму интервалов и гистограмму распределения интервалов относительно начала действия стимула. Если первый тип гистограммы отражает наиболее вероятный интервал между соседними пиковыми разрядами нейрона, то второй тип показывает наиболее вероятную структуру ответной реакции нейрона после нанесения раздражителя на его рецептивное поле (рис. 70). Оба типа гистограмм, вместе взятые, отражают также преобразование ф. а. при нанесении раздражителей, активирующих или тормозящих деятельность нейрона. Некоторые механизмы взаимодействия фоновой и вызванной активности нейрона были изучены А. И. Шаповаловым (1961б, 1962).
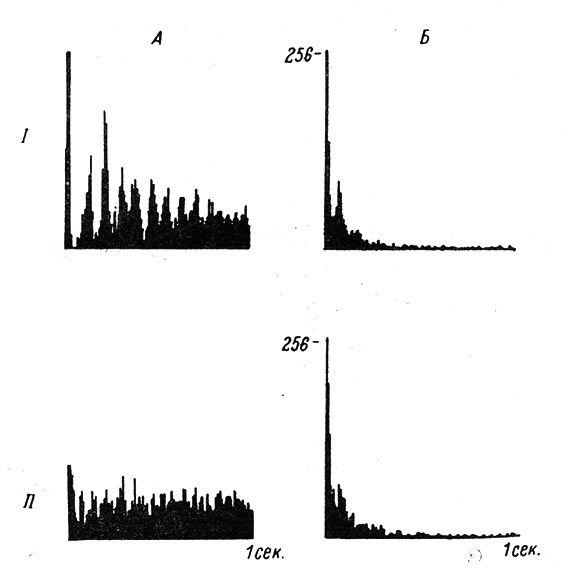
Рис. 70. Два вида анализа межимпульсных интервалов нейрона в слуховой коре кошки. А - интервальная гистограмма при слуховом раздражении, построенная относительно начала раздражения (I); интервальная гистограмма фоновой активности, построенная относительно произвольного начального момента отсчета (II). Б - общая гистограмма распределения межимпульсных интервалов при слуховом раздражении (I) и при фоновой активности (II) нейрона (Grerstein, 1960). Длительность горизонтальной оси - везде 1 сек. По вертикали - число интервалов соответствующей длительности
При вызванной активности уменьшается вариабильность межимпульсного интервала, например, для таламических нейронов с 93 ± 2.9% до 52 ± 1.7% (Werner a. Mountcastle, 1963). Было высказано предположение, что это свойство вызванной активности связано с механизмом дифференциальной чувствительности, так как оказалось, что с увеличением интенсивности реакции еще больше уменьшается ее коэффициент вариации, т. е. еще плотнее распределение пиков во времени (рис. 71). Если бы вариабильность не изменялась, то при нелинейных соотношениях, чрезвычайно характерных для передаточных функций нейрона, невозможно было бы обеспечить различение близких градаций вызванной реакции, когда сама величина межимпульсного интервала изменяется очень незначительно (см., например, рис. 71, А, 2-й интервал Δθ).
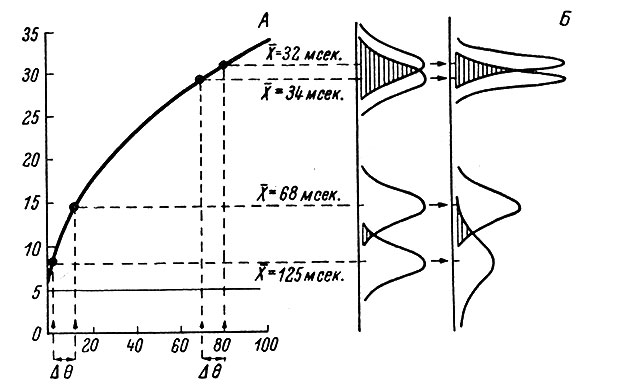
Рис. 71. Механизм различения градаций сигнала в таламическом нейроне 'положения'. (Werner a. Mountcastle, 1963). А - реакция при различных углах сгибания конечности. По оси абсцисс - угол сгибания конечности, в градусах; по оси ординат - вызванная активность, в имп./сек. Справа от графика приведены средние значения межимпульсного интервала для некоторых углов сгибания конечности при одинаковых Δθ = 10°. Б - два ряда расчетных распределений межимпульсных интервалов, относящихся к средним х, приведенным на А. Справа - увеличение угла сгибания на Δθ° в двух случаях приводит к различному уменьшению межимпульсного интервала, в то время как его стандартное отклонение в обоих случаях уменьшается вдвое. Слева - распределение при постоянном стандартном отклонении, равном 30 мсек
Автокорреляционный анализ вызванной активности выявил периодические ее колебания. Так, у таламических нейронов "положения" периодические усиления и ослабления вызванной активности происходили с частотой от 0.09 до 1.32 периода в 1 сек., причем эти колебания оказались не случайными, что подтверждается статистической проверкой методом Монте-Карло (рис. 72; Werner a. Mountcastle, 1963).
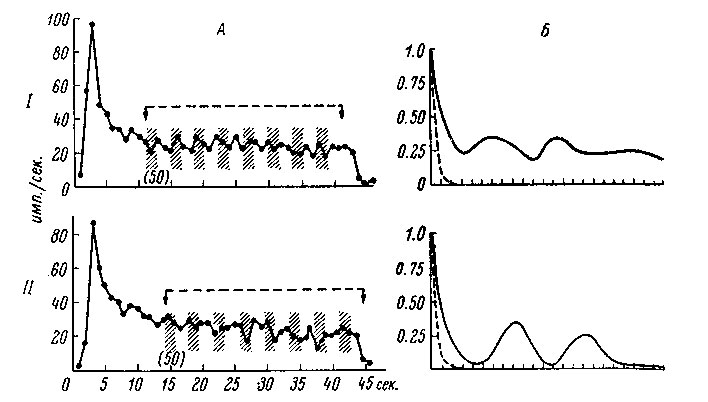
Рис. 72. Определение вариабильности вызванной активности таламического нейрона. (Werner a. Mountcastle, 1963). На I и II показаны реакции на одинаковые сгибания сустава под углом 80°. По оси абсцисс: на А - время, в сек., на Б - сдвиг по 200 мсек. По оси ординат: на А - частота вызванной активности, в имп./сек., на Б - величина автокорреляционной функции. На А стрелками показан период анализа и размеры отдельных отрезков, автокорреляционная функция которых после случайного перемешивания представлена на Б прерывистой линией. Б - автокорреляционные функции вызванной активности. При первом раздражении спектральная плотность (не показано) имеет пик на частоте 0.68 периода в 1 сек., при втором - 0.78 периода в 1 сек
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'