III. Фоновая импульсная активность и электроэнцефалограмма
Специальный интерес представляет изучение связи фоновой импульсации отдельных нейронов с суммарными биоэлектрическими потенциалами.
В настоящее время установлено, что суммарные электрические процессы типа ЭЭГ, ЭКоГ вызванного потенциала и другие имеют иную природу, чем пиковая активность в соме или аксоне нейрона (Li, McLennan a. Jasper, 1952; Li a. Jasper, 1953; Пурпура, 1959, и др.). ЭЭГ или ЭКоГ является результатом суммации постсинаптических потенциалов и колебаний мембранных потенциалов в дендритах и телах корковых нейронов. Совершенно очевидно, что не всякие колебания мембранного потенциала доводят поляризацию нейрона до критического уровня и сопровождаются появлением ПД, поскольку ПСП могут быть подпороговыми, а также не только деполяризовать, но и гиперполяризовывать мембрану нейрона.
Ли и Джаспер (1953), Ли, Куллен и Джаспер (Li, Cullen, a. Jasper, 1956a, 1956b) и другие отметили очень слабую связь между пиковой активностью и поверхностной ЭКоГ. Они наблюдали лишь единичные клетки, пиковые разряды которых появляются во время определенных волн ЭКоГ или их фаз. Однако в случаях выраженной синхронизации активности в отдельных участках мозговой ткани чаще выявляются более определенные соотношения. Так, например, пиковая активность отдельных нейронов в фокусе вызванного потенциала более тесным образом связана с отдельными его фазами (Gerstein, 1960; Джавришвили, 1963, и др.); то же наблюдается при развитии реакции вовлечения в коре (Li, Cullen a. Jasper, 1956a, 1956b, и др.), во время стрихнинных разрядов (Thomas et al., 1955, и др.), при введении судорожных средств и развитии аноксии (Li a. Jasper, 1953), в эпилептическом очаге (Ward, 1961; Matsumoto a. Ajmone Marsan, 1964, и др.) и т. д.
П. К. Анохин (1964) отметил наиболее выраженные изменения пиковой активности в моменты смены частоты и амплитуды волн электроэнцефалограммы. Однако и в этих случаях связь пиковой активности с определенными волнами ЭКоГ остается разнообразной и изменчивой. Возможно, что причиной этого является флуктуирующий характер фоновой и вызванной активности, а также постоянные колебания возбудимости нейронов (Вартанян и Василевский, 1963 б).
Особенно трудна оценка соотношений пиковой активности с медленными волнами при развитии процессов десинхронизации или при выраженной депрессии мозговой активности (Джаспер, Риччи и Доун, 1958, и др.).
Изменчивый и разнообразный характер медленных электрических процессов, а также большая вариабильность активности отдельных нейронов позволяют считать, что для исследования временных соотношений между двумя этими процессами наиболее целесообразно применять статистические методы анализа. В этом направлении уже получены некоторые результаты. Так, Грин и сотрудники (Green et al., 1960), используя специальное электронное устройство, позволяющее регистрировать пиковые потенциалы в определенные фазы гипокампального тета - ритма, обнаружили у большинства нейронов наличие более или менее выраженной тенденции к появлению пиков в определенные фазы медленной электрической активности (рис. 62), причем эти особенности распределения пиков по фазам не выявлялись при обычном визуальном анализе осциллограмм. Дальнейшая разработка этого вопроса имела место в исследовании Н. Н. Василевского (1965б) по анализу соотношений между поверхностными волнами ЭКоГ и коэффициентом асимметрии (к. а.) отдельных нейронов на основе статистических приемов. В частности, автора специально интересовал вопрос, имеются ли особенности этих связей у нейронов, расположенных на различной глубине в коре и в зависимости от уровня основной ф. а.

Рис. 62. Распределение разрядов отдельных нейронов гиппокампа относительно вектора ЭЭГ, вращающегося как показано на диаграмме справа вверху. (Green et al., 1960). Каждая точка диаграммы отражает момент появления одиночного импульса нейрона. Положение точек в том или ином квадранте определяется фазой ЭЭГ в момент разряда нейрона, а расстояние от точки до центра диаграммы - амплитудой ЭЭГ в данный момент. Диаграммы получены при многократном наложении отдельных циклов
Для разрешения поставленных задач с помощью микроэлектродов диаметром 1-4 мм отводилась импульсная активность от нейронов в соматосенсорной зоне коры больших полушарий кошки. Вблизи места погружения микроэлектрода одновременно с ПД нейрона регистрировалась поверхностная монополярная ЭКоГ. Для анализа статистических соотношений был избран метод разделения электрокортикограммы на фазы по А. А. Генкину (1963). Исследовались 5-10-секундные отрезки осциллограмм, причем в каждой секунде выделялось 60 интервалов. Подсчитывалось общее число пиков, приходящееся отдельно на восходящие и нисходящие фазы ЭКоГ. Отношение большей суммы к меньшей принималось за к. а. распределения пиков по фазам ЭКоГ. В каждом случае высчитывалась достоверность наблюдаемого распределения по t-критерию при данном количестве зарегистрированных пиков и числе выделенных фаз.
Из 127 изученных нейронов только у 12 наблюдалась явная визуальная связь импульсной активности с фазами ЭКоГ, причем 7 из них располагалось на глубине до 1 мм и 5 - больше 1 мм. В остальных случаях визуальный анализ не мог дать определенного результата. Вычисление к. а. показало, что в среднем его значение у отдельных нейронов убывает по мере удаления от поверхности коры (рис. 63). Индивидуальные значения к. а. колебались от 1 до 4. Достоверные величины к. а. наблюдались у 38% нейронов (р = 0.05), причем среднее его значение у этой группы клеток оказалось равным 2.08 ± 0.16. Значимые величины к. а. в основном наблюдались у поверхностно расположенных нейронов, хотя в отдельных случаях, но очень редко (0.8%) наблюдались нейроны со значимым распределением пиков по фазам и в глубоких слоях коры (рис. 63).
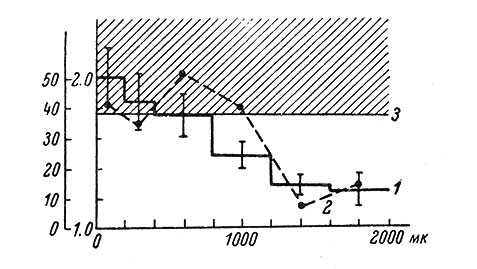
Рис. 63. Средние значения коэффициента асимметрии у нейронов, находящихся на различной глубине в коре. (Ориг.). 1 - средняя величина коэффициента асимметрии; 2 - процент нейронов, имеющих достоверное распределение пиков по фазам ЭКоГ; 3 - нижняя граница достоверности средней величины коэффициента асимметрии при р = 0.05. Вертикальные отрезки - величина стандартного отклонения. По оси абсцисс - глубина от поверхности, в мк; по оси ординат - величина коэффициента асимметрии и процент нейронов со значимым распределением пиковых потенциалов по фазам ЭКоГ
Нейроны с достоверным распределением импульсов по фазам ЭКоГ встречаются действительно чаще в поверхностных слоях и преимущественно на глубине до 1 мм, главным образом в 3-м и 4-м слоях. Эти слои, как известно, являются наиболее физиологически активными (Ройтбак, 1955).
Кроме того, было показано, что средняя величина к. а. зависит и от частоты фоновой активности (табл. 11).

Таблица 11. Зависимость между уровнем фоновой активности корковых нейронов и средней величиной к. а.
Таким образом, исследование Н. Н. Василевского показало, что в каждом слое коры имеется сравнительно небольшая часть нейронов, фоновая пиковая активность которых проявляется преимущественно в определенные фазы (восходящие или нисходящие) медленной электрической активности. Тот факт, что нейроны с достоверным распределением пиков по фазам ЭКоГ встречались чаще в поверхностных слоях коры, по-видимому, связан с особенностями формирования ЭКоГ, в которой преимущественно отражается активность нейронов более поверхностно расположенных слоев коры больших полушарий (Ройтбак, 1955; Amassian, 1961, и др.).
Обнаруженные соотношения между величиной к. а. и частотой ф. а., нам кажется, наиболее рационально объяснить различиями в поляризации клеточной мембраны. Как уже отмечалось выше, Сасаки и Ока (Sasaki a. Oka, 1963) на мотонейронах глазодвигательного ядра показали, что особенности фоновой, а также вызванной активности тонических и фазических мотонейорнов объясняются различиями как поляризации клеточной мембраны, так и аккомодационных свойств и амплитуды постсинаптических потенциалов. Было отмечено, что при высоком уровне ф. а. у так называемых тонических мотонейронов уровень МП весьма близок к критическому. При этом незначительные по амплитуде ПСП сопровождаются импульсными разрядами. Напротив, у фазических нейронов с редкой ф. а. только значительные ПСП приводят к разряду клетки в аксон.
Эти данные позволяют полагать, что группа клеток с низкой фоновой импульсной активностью должна проявлять более тесную связь с отдельными фазами ЭКоГ, поскольку ПСП таких нейронов принимает большее участие в формировании поверхностной ЭКоГ. У первой группы клеток (с высоким уровнем ф. а.) ПСП принимает меньшее участие в образовании отдельных волн медленной электрической активности. Наиболее вероятно, что особенности поляризации в этой группе нейронов находят отражение в уровне постоянного потенциала коры больших полушарий (Аладжалова, 1962; Мнухина, 1963, и др.).
В последнее время Фром и Бонд (Fromm a. Bond, 1964) показали, что часть корковых нейронов разряжается преимущественно во время положительной фазы медленного потенциала и тормозится во время отрицательной полуволны. Этот результат понятен в связи с данными Филлипса (Phillips, 1959), который показал, что гиперполяризация сомы пирамидных нейронов коры совпадает с появлением негативных волн на ее поверхности. По аналогии с этими данными можно также предположить, что деполяризации сомы (ВПСП) будет соответствовать положительное колебание постоянного потенциала коры. Однако эти соотношения, по-видимому, наблюдаются только при определенных состояниях коры, поскольку при чрезмерной деполяризации сомы и дендритов, когда развивается состояние, подобное катодической депрессии, импульсная активность нейрона тормозится. Поэтому в каждом конкретном случае вопрос о происхождении торможения импульсной активности нейрона может быть решен только на основе результатов внутриклеточных отведений.
Н. Н. Василевским и В. В. Усовым была сделана попытка оценить временные соотношения между фоновой активностью отдельных нейронов коры и поверхностной и глубокой ЭКоГ путем определения взаимокорреляционной функции с помощью электронно-вычислительной машины. На рис. 64 показан пример взаимокорреляционной функции пиковой активности нейрона с поверхностной и глубокой ЭКоГ. Отмечаются небольшие колебания коэффициента корреляции в положительной и отрицательной области для взаимокорреляционной функции импульсной активности нейрона как с поверхностной, так и с глубокой электрокортикограммой (рис. 64 и 65). Весь вид взаимокорреляционной функции, так же как и корреляция в нулевой точке, сильно варьирует в зависимости от начала отсчета. Эти вариации превышают вариации, зависящие от объема выборки, что свидетельствует о нестационарности процесса.
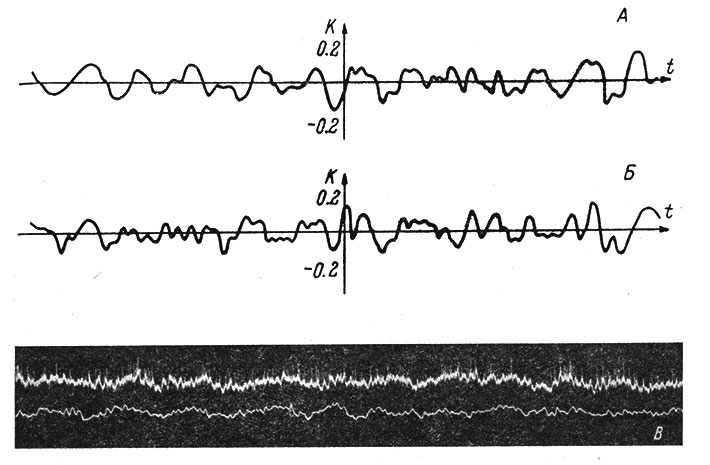
Рис. 64. Взаимокорреляционные функции импульсной активности коркового нейрона кролика с поверхностной и глубокой ЭКоГ. (Ориг.). А - ЭКоГ, снятая макроэлектродом вблизи места погружения микроэлектрода; Б - глубокая ЭКоГ, зарегистрированная через микроэлектрод; В - оригинальная осциллограмма пиковой активности нейрона (вверху) и поверхностной ЭКоГ (внизу). Отметка времени 1/20 сек. Нижняя полоса пропускаемой частоты в обоих отведениях равнялась 1 гц, верхняя для ЭКоГ - 150 гц, для микроэлектродного канала - 8000 гц
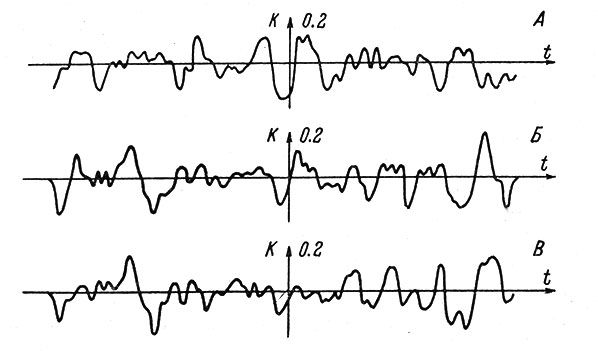
Рис. 65. Зависимость формы взаимокорреляционной функции, соответствующей рис. 64, А, от сдвига начала отсчета на исследуемых кривых. (Ориг.). А - длина реализации 5 сек. (начало отсчета соответствует началу на рис. 64, А). Б - сдвиг на 3.3 сек. В - сдвиг на 5 сек. относительно начала отсчета на А
Этот вывод позволяет считать, что для оценки связи подобного рода процессов необходимо разрабатывать специальные статистические приемы, основанные непосредственно на свойствах нестационарных процессов. Поскольку классический статистический прием, каким является взаимокорреляционная функция, оказался малоэффективным, мы полагаем, что непараметрический метод А. А. Генкина, давший более четкие результаты, будет более "чутко" реагировать на наличие связи между двумя процессами, даже в случаях ее нестационарности. В последнее время нестационарная природа физиологических функций настойчиво подчеркивается Г. А. Сергеевым и А. Ф. Романенко (1964).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'