II. Фоновая импульсная активность
Среди разнообразных форм деятельности нейрона особый интерес представляет изучение ПД - импульсной или "пиковой" активности, поскольку она есть результирующая многочисленных влияний, падающих на нейрон, и является выражением его эфферентной функции. В настоящее время принято различать фоновую (иногда именуемую "спонтанной") и вызванную импульсную активность. В многочисленных исследованиях подробно изучены различные параметры фоновой и вызванной активности: частота, распределение пиков во времени, их вариабильность и др.
Выделено несколько типов фоновой и вызванной активности, показаны их особенности в отдельных сенсорных системах и ядрах. Однако физиологическая роль и особенно информационное значение обоих видов активности нейрона во многом остаются неясными. Поскольку разрешение этого вопроса представляет не только значительные методические, но и теоретические трудности, систематизация современных представлений о природе фоновой и вызванной активности представляется целесообразной, так как это позволяет наметить некоторые новые пути дальнейшего анализа.
Под фоновой активностью обычно понимается такая импульсация в нейроне, которая имеет место вне связи с наносимыми раздражениями или в их отсутствие. Ошибочно полностью отождествлять ф. а. со спонтанной, поскольку последняя представляет собой частный случай фоновой импульсации. В центральной и периферической нервной системе, особенно в рецепторах (Гранит, 1956), имеется достаточно много причин, которые могут привести к возникновению истинной формы спонтанной активности, т. е. импульсации, не связанной с информационными процессами и координированной активностью отдельных систем нейронов. Такая импульсация может проявиться, например, при высоком уровне возбудимости рецепторов и отдельных нейронов, особенно в моменты усиления хаотических колебаний ионных потоков через мембрану нейрона, колебаний температуры, интенсивности обмена веществ и т. д. (Pecher, 1939; Вартанян и Василевский, 1963б, и др.). Такую же роль могут играть случайные выделения отдельных квантов передатчика в синапсах (Fatt a. Katz, 1952), поскольку в отдельных случаях для достижения критического уровня разряда вполне достаточно несколько квантов медиатора (Blackman et al., 1963). Таким образом, возможно, что ф. а. состоит из двух компонентов, один из которых отражает циклические координированные процессы в нервной ткани, а другой является истинной формой спонтанной импульсации.
П. Г. Костюк (1965) различает эндо- и экзогенные причины фоновой активности. Однако в каждом конкретном случае трудно определить, какими именно механизмами обусловлена импульсация. Как будет показано ниже, интересные результаты дало изучение связи между особенностями фоновой активности и функциональной характеристикой тех или иных ядер, их функциональным состоянием, уровнем возбудимости и другими параметрами нервной системы.
Ф. а. имеет разнообразное значение. Гранит (1955) определяет ее как важнейшую часть функционального плана нервной системы, которая, по его мнению, с одной стороны, отражает деятельность тонических механизмов ц. н. с., а с другой стороны, возможно, является показателем периодической смены активности отдельных единиц нервной ткани.
Некоторые авторы (Wall, 1959) рассматривают ф. а. как своеобразный шум в нервной системе. Другие расценивают ее как следовое отражение пространственно-временной активности нейронных сетей, связанной с существованием реверберирующих цепочек нейронов (Lorente de No, 1934). Такого рода цепочки нейронов несомненно имеют место и в отдельных случаях являются причиной проявления длительной активности нейрона (сети) после нанесенных раздражений (Лебедев, 1962; Фессар, 1964, и др.).
По мнению Гранита (1955) и Юнга (Jung, 1961), функциональное значение ф. а. может заключаться и в том, что она обеспечивает возможность реагирования системы и отдельных нейронов в двух направлениях: как в сторону усиления, так и в сторону торможения активности. Система нейронов, лишенная ф. а., способна активироваться только в одном направлении. Кроме того, как отмечает Юнг, наличие ф. а. позволяет избежать фиксированных порогов, обеспечивая при этом высокий уровень дифференциальной чувствительности и гибкость системы в целом.
Имеются данные о связи фоновой активности с функциональным состоянием ц. н. с. Еще Н. Е. Введенский (1901), Н. В. Голиков (1950), Д. Н. Насонов (1959) и многие другие непосредственно связывали уровень ф. а. с состоянием возбудимости. Эта точка зрения подтверждается тем, что ф. а. центральных нейронов резко подавляется при любых видах наркоза (Frank a. Fuortes, 1955, 1956а, 1956b; Hunt a. Kuno, 1959a, 1959b; Костюк, 1960, и др.). Последнее особенно проявляется на промежуточных нейронах (Лебедев, 1952; Шаповалов, 1964, и др.). Связь ф. а. нейронов с функциональным состоянием ц. н. с. подтверждают также данные об особенностях импульсации нейронов в эпилептическом очаге.
Мартин и Бранч (Martin a. Branch, 1958) считают, что в эпилептическом очаге резко увеличивается число клеток с высокочастотной активностью. Такой тип активности клетки встречается и в нормальной ткани, но значительно реже. Шмидт и сотрудники (Schmidt et al., 1959) полагают, что повышенная активность "эпилептического" нейрона связана с длительной и устойчивой деполяризацией дендритов и особенно сомы нервных клеток, которая в отдельных случаях может быть обусловлена механической деформацией мембраны нейрона (например, рубцовой тканью; Ward, 1961).
Отмечая высокую частоту ф. а., необходимо остановиться на одном из возможных артефактов. При введении микроэлектрода в толщу нервной ткани как при вне-, так и особенно при внутриклеточной регистрации в первый момент обнаружения нейрона отмечается более высокая частота ф. а., которая, однако, в течение нескольких минут снижается и стабилизируется на каком-то уровне (иногда пропадает полностью). Очевидно, в этих случаях проявляется раздражающее действие погружаемого в ткань микроэлектрода.
Юнг (Jung, 1961) полагает, что длительно наблюдаемые разряды нейронов коры с частотой 800-1000 имп./сек. всегда являются выражением патологических процессов, которые либо связаны с повреждением нейрона микроэлектродом, либо отражают эпилептические разряды. По его мнению, в нормальных условиях импульсация с частотой 400-500 имп./сек. может наблюдаться только в течение очень коротких промежутков времени.
Выявлена связь между характером ф. а. и уровнем развития ц. н. с. Так, в сравнительно-физиологическом ряду: аксолотль - амблистома - лягушка - агама обнаружена неодинаковая величина (средняя частота) ф. а. нейронов зрительной покрышки, а также различное процентное содержание нейронов, обладающих такой активностью. Более редкий ритм и больший процент "молчащих" нейронов характерен для низших животных этого ряда. Значительная часть нейронов по сравнению с теплокровными (кошки, кролики и др.) не имеет ф. а. и проявляет импульсацию только при нанесении раздражения (Смирнов и Мантейфель, 1962; Смирнов и Мазурская, 1963; Смирнов, 1963а, 1963б).
А. Б. Коган (1964) показал, что в эволюционных рядах отмечается также закономерное нарастание аритмичности импульсации, что отражает усложнение межнейронных связей и является одним из условий их оптимального функционирования (Меницкий, 1964).
По данным Л. Д. Карпенко (1964), в подглоточном ганглии виноградной улитки число нейронов, имеющих ритмический тип ф. а., в 3 раза больше, чем нейронов с аритмической импульсацией. Если обратиться к характеристике нейронов спинного мозга кошки (Лебедев, 1962), то ритмичная активность наблюдается только в 21.3% всех нейронов, имеющих ф. а.
Некоторые экспериментальные данные дают основание также связывать уровень ф. а. нейронов определенных отделов ц. н. с. с особенностями функциональной организации их афферентных систем. Так, нейроны 3-го поля соматосенсорной коры обезьян, имеющие связи преимущественно с кожными рецепторами, обладают гораздо более высокой ф. а., чем нейроны 1-го и 2-го поля той же корковой зоны, связанные главным образом с проприорецепторами (Mountcastle a. Powell, 1959a, 1959b).
Различными оказались уровни ф. а. нейронов различных типов в соматосенсорной коре (SI и SII) кошек и кроликов (рис. 59). В частности, у кошек наибольшей частотой фоновой импульсации обладают нейроны с распространенными рецептивными полями (28.2 ± 2.2 имп./сек.), имеющие обширные полимодальные и гетеротопические рецептивные поля. Нейроны с локальными одномодальными рецептивными полями имеют более низкую ф. а. (19.1 ± 1.8 имп./сек.). Наименьший уровень ф. а. характерен для неактивирующихся нейронов (8.9 ± 2.3 имп./сек.), у которых не удалось обнаружить рецептивные поля в пределах кожного и мышечного анализаторов (Василевский, 1965а).
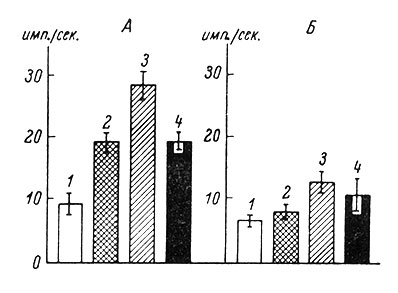
Рис. 59. Средние уровни фоновой активности у отдельных типов нейронов в соматосенсорной области коры кошек (А) и кроликов (Б). (Василевский, 1965). 1 - нейроны без выявленного рецептивного поля; 2 - нейроны с локальным рецептивным полем; 3 - нейроны с распространенным рецептивным полем; 4 - средний уровень для всех типов нейронов. Вертикальные отрезки - величина стандартного отклонения
Брукс (Brooks, 1960) также отметил низкий уровень фоновой активности у нейронов с локальными рецептивными полями.
Высокий уровень ф. а. корковых нейронов с распространенным рецептивным полем можно с полным основанием связать с обилием афферентных путей, конвергирующих на этих нейронах, поскольку в рецепторных образованиях всегда имеет место некоторый уровень ф. а. (Гранит, 1956).
Эти нейроны соматосенсорной зоны по уровню ф. а. напоминают нейроны ретикулярной формации (Amassian a. De Vito, 1954; Лиманский, 1961, 1963, и др.). Сасаки и Ока (Sasaki a. Ока, 1963; Sasaki, 1963) отметили высокий уровень ф. а. у тонических и значительно более редкую импульсацию у фазических мотонейронов.
Временное распределение, или так называемая структура фоновой импульсной активности, чрезвычайно разнообразно. Целесообразно различать единичную, пачковую и групповую импульсацию (рис. 60).
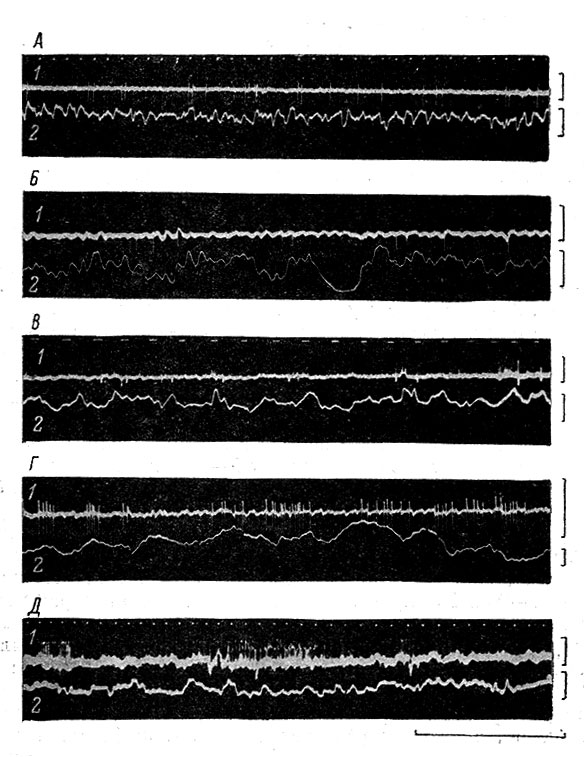
Рис. 60. Различные типы фоновой активности у отдельных нейронов в коре кошек (А, В, Д) и кроликов (Б, Г). (Ориг.). А, Б - единичный тип; В - пачковая, Г, Д - групповая активность. 1 - внеклеточная электронейронограмма (ЭНГ); 2 - поверхностная ЭКоГ. Калибровка для ЭНГ - 5 мв, для ЭКоГ - 100 мкв; время - 1 сек
Единичная активность представляет собой отдельные импульсы, возникающие в различной последовательности. У большинства нейронов имеют место большие вариации в длительности межимпульсных интервалов, особенно при редкой средней частоте. Однако в отдельных случаях наблюдается довольно строгая ритмичность фоновой импульсации.
Пачковый тип ф. а. характеризуется тем, что отдельные импульсы группируются в короткие высокочастотные серии разрядов - пачки. У корковых нейронов такие пачки содержат, как правило, 2-3 импульса; у спинальных промежуточных нейронов число импульсов в серии может достигать 5-7. Интервалы между импульсами в пачке обычно малы (1-4 мсек.) и варьируют незначительно, несколько нарастая от предыдущего к последующему. В то же время межпачковые интервалы колеблются в довольно широких пределах.
При групповом виде активности в каждом периоде деятельности нейрона может насчитываться от 5-7 до нескольких десятков отдельных пиковых потенциалов, разделенных между собой более длительными интервалами, чем при пачковой активности. Нередко между пачками и группами могут возникать и одиночные разряды.
Г. Смит и Д. Смит (G. Smith a. D. Smith, 1964) выделили два типа ф. а., один из которых представлен высокочастотными (порядка 50-60 в 1 сек.) разрядами, что позволило авторам назвать этот тип активности "вспышкой". Второй тип активности также имел два ритма, смена которых один на другой, однако, была закономерной. В связи с этим авторы полагают, что существуют два независимых механизма генерации разрядов в нейронах, один из которых определяет редкую активность, другой - быструю.
Конечно, описанные разновидности ф. а. представляют собой лишь наиболее отчетливые ее варианты. Обычно же можно наблюдать разнообразные смешанные и переходные формы фоновой импульсации.
В основе фоновой импульсной активности могут лежать периодические колебания возбудимости электрогенной мембраны (Grundfest, 1957), а также временные и амплитудные особенности деполяризационных процессов в соме нейрона (Буллок, 1959; Костюк, 1965, и др.). На основании имеющихся экспериментальных материалов можно полагать, что единичный тип ф. а. связан с короткими пороговыми деполяризационными процессами. Пачковая активность может возникать при более интенсивной, но все же относительно кратковременной деполяризации. Групповая активность, по-видимому, возникает при длительной и достаточно интенсивной деполяризации. Во всяком случае, такой характер активности можно наблюдать при длительных сдвигах уровня мембранного потенциала при искусственной деполяризации нейрона электрическим током (Frank a. Fuortes, 1956b; Костюк и Шаповалов, 1960), а также при некоторых видах синаптической стимуляции (Koizumi et al., 1959; Sasaki et al., 1960; Вартанян и Василевский, 1963a; Вартанян и Василевский, 1964а, и др.). Кроме того, после пачкового разряда нейрона может наблюдаться фаза активного торможения нейрона (Martin a. Branch, 1958).
Ранее отмеченные особенности ф. а. тонических мотонейронов по сравнению с фазическими Сасаки и Ока (Sasaki a. Oka, 1963; Sasaki, 1963) объясняют низкой аккомодацией мембраны, а также близостью ее МП к критическому уровню разряда. Кроме того, у тонических нейронов слабо выражено стабилизирующее влияние возвратного торможения.
Могут ли отражаться морфологические особенности отдельных нейронов на характере их фоновой и вызванной активности? Некоторые данные показывают, что имеется вполне определенная связь. Л. Д. Карпенко (1964) показала, что в висцеральном ганглии улитки ритмичные типы ф. а. преимущественно встречаются у крупных клеток, неритмичные - у средних и мелких. Генеман (Неnnemann, 1957, 1965) считает, что уровень возбудимости может также определяться размерами нейронов. Некоторые расчеты позволяют понять эту связь. Поскольку у нейронов небольших размеров поверхность соматической мембраны мала, то при активации синапсов плотность деполяризующего или гиперполяризующего тока будет значительно больше, чем у крупных нейронов при возбуждении такого же количества синаптических окончаний. Поэтому у мелких нейронов следует ожидать более крутого нарастания постсинаптических потенциалов в области сомы нейрона, а также более интенсивную ее общую деполяризацию или гиперполяризацию. Так, Экклс и сотрудники (J. С. Eccles, R. M. Eccles, Iggo a. Lundberg, 1961) показали, что у клеток Реншоу, которые, по данным В. И. Пилипенко (1962), являются сравнительно мелкими нейронами, ВПСП может достигать десятков милливольт. При этом ответная реакция клетки представляется в виде высокочастотных пачек пиковых потенциалов. Вследствие такой интенсивной деполяризации нередко отмечается кратковременная инактивация натриевого механизма (катодическая депрессия), что проявляется в изменении межимпульсных интервалов в начале пачкового разряда (см., например, осциллограммы на рис. 66, где на В, Г, Д и Е имеется выпадение очередного пика после первого разряда).
Возможно, что именно эти особенности строения и проявления постсинаптических потенциалов и определяют исключительную реактивность большинства вставочных нейронов, имеющих небольшие размеры тела клетки. Не исключено, что размеры нейрона являются своеобразным регулятором активности нейрона.
С количественной стороны ф. а. следует рассматривать как случайный процесс. В связи с этим для его описания целесообразно привлечь некоторые статистические методы. Поскольку в большинстве случаев ф. а. не является строго периодической функцией, то прежде всего необходимо количественно оценить степень ее вариабильности и наиболее вероятную частоту следования импульсов. Эти показатели могут быть определены при изучении гистограмм распределения межимпульсных интервалов. Гистограммный метод анализа импульсной активности был предложен Герштейном (Gerstein a. Kiang, 1960; Gerstein, 1960) и в настоящее время применяется многими исследователями (Преображенский и Яровицкий, 1962, 1963; Чораян, 1962, 1963а, 1963б; Карпенко, 1964, и др.).
Характер гистограмм, построенных на достаточно большом числе интервалов, исключительно точно отражает структуру ф. а. Так, при ритмической активности любой частоты распределение интервалов будет плотнее, чем при менее регулярной активности. Вычисление среднеквадратичного отклонения (σ), а также коэффициента вариации (υ = σ/Δt ⋅ 100%) позволяет количественно оценить величину разброса и флуктуации межимпульсного интервала. Например, Вернер и Моунткастл (Werner a. Mountcastle, 1963) нашли среднюю величину коэффициента вариации для таламических нейронов равной 93 ± 2.9%, а также определили зависимость между величиной межимпульсного интервала и его стандартным отклонением (рис. 61).
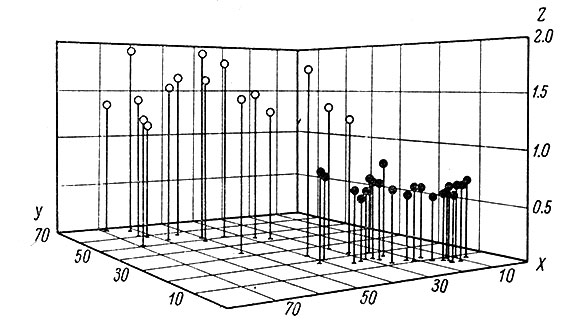
Рис. 61. Соотношения между средним интервалом, в мсек. (ось X), стандартным отклонением (ось Y) и коэффициентом регрессии стандартного отклонения на средний интервал (ось Z) для фоновой (белые кружки) и вызванной (черные кружки) активности у нейронов вентробазального комплекса таламуса. (Werner a. Mountcastle, 1963)
При сложных формах импульсной активности, например при сочетании единичной регулярной или нерегулярной импульсации с пачковой или групповой активностью, а также при "чистой" пачковой и групповой активности, характер распределения будет либо двувершинным, либо с одной вершиной и плато. В этом случае первая вершина будет отражать вариации межимпульсного интервала внутри пачек и групп, а вторая вершина или плато - межпачковые или межгрупповые интервалы, либо интервалы промежуточной единичной активности.
Левик, Бишоп, Вильямc и Лампард (Levick, Bishop, Williams a. Lampard, 1961), используя анализатор распределения вероятностей, обычно применяемый в ядерной физике, показали различные типы распределения интервалов между импульсами по ходу зрительной системы. В сетчатке интервалы распределялись по закону Гаусса, в коленчатом теле - по закону Пуассона, в протектальном ядре - с вершиной и плато.
В исследовании случайных функций особенно важное значение приобретает методика обнаружения периодичности. Некоторые статистические приемы, - такие как анализ Фурье и стандартные статистические методы, - оказывались удовлетворительными только для чисто периодических функций (Мерсер, 1961). Если же периодический сигнал замаскирован случайными компонентами, то эффективность обычных методов оказывается недостаточной. В разрешении проблемы выделения сигнала из шумов наиболее эффективными оказались методы статистической теории связи (Винер, 1948).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'