V. Возбудимость и тонус нервных "центров"
До сих пор речь шла о механизмах регуляции возбудимости одиночных нервных клеток. Теперь целесообразно вкратце рассмотреть, как свойства возбудимости отдельных нейронов проявляются в деятельности однородной совокупности нейронов - нервного "центра". Проще всего это сделать на примере совокупности мотонейронов ("мотонейронного пула") при ее моносинаптической активации.
При раздражении волокон группы I, А афферентного мышечного нерва сверхпороговым одиночным стимулом в МН, иннервирующих соответствующие мышцы,* возникают моносинаптические ВПСП. В зависимости от числа синаптических контактов и уровня поляризации часть МН деполяризуется до порогового уровня, и в них происходит импульсный разряд (ПД); другая, обычно более значительная часть МН этого пула не разряжается, но на время развития моносинаптического ВПСП возбудимость этих клеток повышается. Эти МН составляют так называемую подпороговую зону нервного "центра".
* (На такой раздражающий стимул реагирует МН мышцы, нерв которой раздражается, и непосредственных мышц-синергистов, действующих на тот же сустав.)
Точными количественными исследованиями Д. Ллойда было показано, что подпороговая зона нарастает при усилении пресинаптического раздражения гораздо круче, чем разряжающая зона (рис. 55), причем при любой интенсивности раздражения подпорогово возбужденных нейронов всегда больше, чем отвечающих импульсной активностью (Lloyd, 1943).
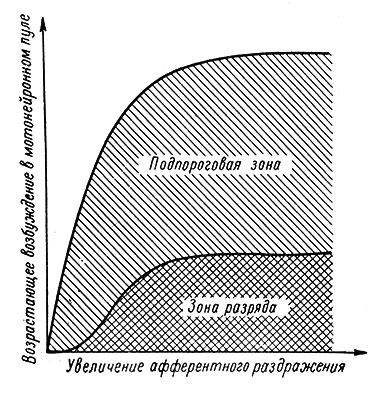
Рис. 55. Полусхематическое изображение соотношений между размерами разряжающейся и подпороговой зон при увеличении силы афферентного раздражения. (Lloyd, 1943)
Эти данные были получены с помощью записи моносинаптических рефлекторных электрических ответов с передних корешков люмбального отдела спинного мозга. Величина переднекорешковых потенциалов отражает число разряжающихся МН, поскольку эти потенциалы формируются в результате суммации импульсов отдельных МН. "Молчащие", но обладающие повышенной возбудимостью МН выявлялись с помощью методики парных импульсов, которая считается теперь классической.
Если разделить задний корешок на две веточки или использовать нервы двух мышц-синергистов и наносить раздельные электрические раздражения на каждый из афферентных источников сначала порознь, а потом одновременно, то оказывается, что во втором случае рефлекторный ответ, записываемый с переднего корешка, значительно больше суммы рефлекторных ответов, получаемых при раздельном раздражении сначала одного, а потом другого афферентного источника.
В настоящее время благодаря микроэлектродной технике совершенно точно выяснено, что происходит при этом в МН, но и до развития этой техники на основании косвенных данных причины этого феномена были раскрыты Ч. Шеррингтоном и его сотрудниками (Sherrington, 1925; Крид, Денни-Броун, Икклс, Лиддел и Шеррингтон, 1935).
В МН подпороговой зоны, как уже говорилось, в ответ на моносинаптический стимул возникает ВПСП, не сопровождающийся развитием ПД. Если одновременно или через небольшой промежуток времени к нему поступает второе моносинаптическое раздражение по другому афферентному пути (например, от нерва мышцы-синергиста), то в нем возникает дополнительная волна ВПСП, которая суммируется с первой и доводит деполяризацию до порогового уровня, вызывая развитие импульса. Поскольку разные афферентные источники конвергируют на одних и тех же МН, то подпороговые зоны МН, возникающие при раздражении двух источников, могут перекрываться. При этом будет наблюдаться примерно следующая картина (рис. 56).
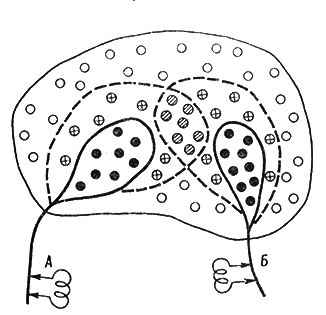
Рис. 56. Схематическое изображение феномена облегчения в нервном 'центре' (Ruch et al., 1963, с модификацией)
Допустим, что раздражение источника А вызывает разряд 8 МН (через кружки) и в 14 МН повышает возбудимость в связи с развитием в них ВПСП (кружки с "+" внутри). Раздражение источника Б вызывает разряд 9 МН ив 15 МН, 7 из которых входит в число подпороговых нейронов первой группы, вызывает только ВПСП. Теперь если наносить раздражения одновременно и на А, и на Б, то разрядится не 17 нейронов, как это должно было бы быть при простой суммации ответов, а 24, потому что 7 нейронов при такой структурно-функциональной организации получает двойную стимуляцию (из А и Б). В этих нейронах произойдет суммация ВПСП, в результате которой они будут генерировать ПД. Поэтому ответ на два раздражения превзойдет по интенсивности сумму ответов на каждое из раздражений. Этот феномен уже давно получил название облегчения.
Изучение облегчения позволило сформулировать представления о разряжающейся и подпороговой зонах нервного "центра". Суммация возбуждений (ВПСП) в нейронах с повышенной возбудимостью приводит к крутому нарастанию эффекта от нескольких (2 и более) раздражений, нанесенных на разные афферентные источники.
Однако разделение "центра" при поступлении одиночного раздражения на разряжающуюся и подпороговую зоны и конвергенция пресинаптических окончаний на одних и тех же нейронах могут приводить к развитию другого феномена, заключающегося в том, что при одновременном раздражении двух афферентов рефлекторный ответ оказывается значительно меньшим, чем сумма ответов на каждое из раздражений. Этот феномен может возникать при несколько ином пространственном распределении конвергирующих пресинаптических окончаний (рис. 57).
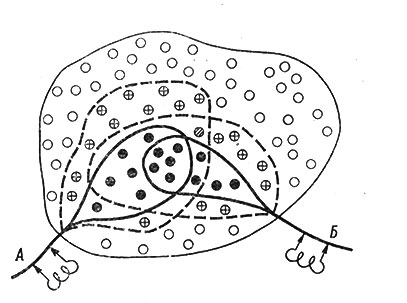
Рис. 57. Схематическое изображение феномена окклюзии в нервном 'центре'. (Крид и др., 1935; Ruch et al., 1963; с модификацией)
На рис. 56 и 57: белые кружки - невозбужденные нейроны; черные кружки - разряжающиеся нейроны; кружки с крестиком - нейроны в состоянии подпорогового возбуждения; заштрихованные кружки - нейроны, которые дают разряд при одновременном раздражении обоих афферентных источников. Показаны только афферентные "входы" центра, эфферентный "выход" не показан. Подробное описание в тексте.
Если при раздражении источника А разряжается 11 нейронов, а при раздражении источника Б - 9, 5 из которых уже вовлеклось в импульсную реакцию при раздражении А, то общее число разряжающихся нейронов будет при одновременном раздражении двух "входов" меньше суммы нейронов, генерирующих импульсы на каждое из раздражений отдельно. При таком распределении пресинаптических окончаний увеличение реакции за счет суммации ВПСП в общих подпороговых нейронах оказывается недостаточным, чтобы покрыть относительное уменьшение числа нейронов, включающихся при одновременном раздражении двух афферентных источников. Так, в схеме, представленной на рис. 57, показан только один совместный для двух источников подпороговый нейрон, в то время как совместных нейронов разряжающейся зоны 5. Другие 2 нейрона, являясь подпороговыми при раздражении источника А, входят в разряжающуюся зону источника Б и также не добавляют числа действующих нейронов при одновременной стимуляции обоих источников. В конечном итоге нанесение раздражения на два источника (А и Б) вызывает импульсные ответы только у 16 нейронов, а не у 20, как это должно было бы быть при простой суммации активности нейронов, разряжающихся на каждое из раздражений (А или Б).
Феномен уменьшенного ответа получил название окклюзии и ничего общего с торможением не имеет. В основе явления лежит общность разряжающихся зон двух или нескольких афферентных источников.
Как же в свете этих данных можно представить себе тонус нервного "центра"?
Не касаясь природы импульсной активности работающих нейронов данного "нервного центра" ("спонтанная" или рефлекторно вызванная активность), можно полагать, что тонус "центра" определяется соотношением числа разряжающихся и "молчащих" нейронов. Если схематически изобразить нервный "центр", состоящий из 50 нейронов, то тонус такого центра гораздо выше, когда импульсная активность имеет место в 25 нейронах из 50 (рис. 58, слева), чем когда разряжается только 11 клеток (рис. 58, справа). Конечно, здесь не учитывается интенсивность (частота) импульсации в разряжающихся нейронах, т. е. их внутренние "тонические" свойства, но этот вопрос частично освещался выше.
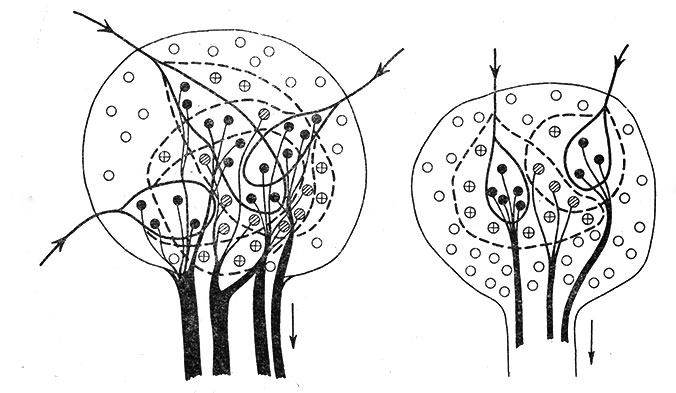
Рис. 58. Схематическое изображение нервных 'центров' с высоким (слева) и низким (справа) тонусом. (Ориг.). Показаны не только афферентные 'входы', но и эфферентный 'выход'. Черная зона 'выхода' показывает относительное количество эфферентных волокон, в которых идет импульсация. Обозначения те же, что на рис. 56. Остальные объяснения в тексте
Памятуя об окклюзии, можно заранее предполагать, что, чем выше тоническая активность центра, т. е. чем большее число нейронов разряжается в данный момент, тем меньше возможности "центра" развивать рефлекторную деятельность в ответ на дополнительные раздражения. Это видно из рис. 58. "Центр" слева находится в состоянии более высокого тонуса, но у него только половина нейронов может включаться в ответ на какие-либо дополнительные раздражения. "Центр" справа обладает более низкой тонической активностью, но у него, так сказать, больше резервов для включения в рефлекторные реакции.
Действительно, широко известно, что центры, которым присущ постоянный тонус, как например центр блуждающего нерва, обладают тем меньшей рефлекторной возбудимостью, чем выше их тоническая активность.
Таковы общие представления о возбудимости и тонусе нервных "центров", которые вытекают из свойств возбудимости отдельных нервных элементов.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'