III. Синаптические механизмы регуляции возбудимости
Не имея в виду касаться зависимости МП от метаболических процессов в нейроне, которые пока изучены еще крайне недостаточно, при условии, что речь будет идти о нейроне, находящемся в оптимальных условиях обмена, можно сказать, что МП зависит главным образом от текущих синаптических влияний.
Большинство центральных нейронов находится под непрерывным воздействием импульсации из различных афферентных* источников. Эта импульсация может вызывать в нейроне как возбуждающие (ВПСП), так и тормозящие (ТПСП) постсинаптические процессы. Благодаря градуальности и суммационной способности постсинаптических потенциалов (ПСП) они взаимодействуют между собой, постоянно изменяя текущий уровень МП. Колебания МП, связанные с непрерывной синаптической "бомбардировкой" и получившие названия "синаптического шума", представлены на рис. 44 (вставка слева внизу). В таком "синаптическом шуме" трудно выделить возбуждающие и тормозящие компоненты. Подобные колебания МП могут иметь место и при возбуждающей, и при тормозящей асинхронной синаптической "бомбардировке", и при взаимодействии обоих видов синаптических влияний. МП отражает суммарный эффект всех постсинаптических процессов в каждый данный момент, и в зависимости от того, доминируют ли возбуждающие или тормозящие воздействия, он то приближается, то удаляется от порогового уровня деполяризации (Kolmodin a. Skoglund, 1960; Вартанян и Василевский, 1963б, и др.).
* (Афферентными здесь условно названы все источники, конвергирующие на данном нейроне. Они могут быть нисходящими и боковыми (от других нейронов той же совокупности).)
Обратно пропорционально величине МП колеблется уровень возбудимости нейрона. В этих условиях даже совершенно идентичные ВПСП могут в одних случаях достигать, а в других случаях не достигать критического уровня и соответственно то вызывать, то не вызывать ПД. Если еще учесть, что сами ВПСП, вызванные идентичными раздражениями, также постоянно флуктуируют по величине (Вартанян и Василевский, 1963б; Вартанян, 1964), то станет очевидным, что порог, а значит, и возбудимость являются величинами не жестко детерминированными, а вероятностными. Конечно, если МП достаточно близок к критическому уровню, то вероятность вызова ПД отдельным синхронизированным ВПСП может достигать 1 даже в случае его сравнительно небольшой величины и при наличии флуктуации.
Таким образом, уровень МП регулирует возбудимость нейрона и оказывает большое влияние на формирование его импульсной активности.
В каждой нейронной совокупности имеются нейроны с высокой и низкой возбудимостью. Например, многие МН одной совокупности обладают низкой возбудимостью (Hunt, 1955a, 1955b; Lloyd a. Mclntyre, 1955; Perl, 1962). При раздражении какого-либо одного афферентного источника только сравнительно небольшая часть МН одной совокупности начинает давать импульсные разряды, остальные не достигают критического уровня деполяризации и остаются "молчащими". Однако возбудимость этих МН тоже повышается, что нетрудно выявить, нанося одновременно парные раздражения на два синергичных афферентных источника (пространственная суммация). "Молчание" большинства МН связано, конечно, и с тем, что количество синаптических окончаний, возбуждающихся при раздражении одного афферентного источника, на разных нейронах неодинаково, а в связи с этим неодинакова и амплитуда ВПСП в разных клетках. Низкий или нулевой индекс разряда* таких нейронов определяется относительно небольшим числом синаптических связей с этим источником, т. е. незначительным представительством данного афферентного источника в синаптическом аппарате нейрона. Разное представительство различных источников в синаптическом аппарате нейрона (соотношение синаптических полей) является одним из важнейших принципов функциональной организации нейронной совокупности, который имеет большое значение в аналитико-интегративной и координационной деятельности нервных "центров".
* (Индекс разряда представляет собой процентное отношение числа импульсных ответов нейрона к числу нанесенных раздражений (Hunt, 1955a, 1955b; Lloyd a. Mclntyre, 1955).)
Характер импульсной активности может меняться не только за счет увеличения амплитуды ВПСП, которая в конечном итоге ограничивается "мощностью" создающего его синаптического поля, но и за счет более или менее устойчивых сдвигов МП при активации других синаптических полей, в том числе формируемых нисходящими афферентными источниками, передающими импульсацию из высших отделов центральной нервной системы (ц. н. с). Другими словами, импульсная активность может регулироваться временными изменениями возбудимости, возникающими в результате дополнительных синаптических влияний. Такие временные сдвиги МП являются оперативным механизмом регуляции возбудимости и пропускной способности нервных клеток.
В основе изменений импульсной активности при устойчивых сдвигах МП лежит по существу пространственная суммация. Известно, что изменение МП нейрона за счет постсинаптических процессов, вызванных активацией одного из афферентных источников, не изменяет ПСП, вызванных с другого источника (Curtis a. Eccles, 1960). Благодаря этому создаются условия для облегчения возникновения импульса, если оба постсинаптических процесса имеют возбуждающий (деполяризационный) характер. И напротив, если один из постсинаптических процессов носит тормозящий (гиперполяризационный) характер, то он препятствует развитию импульса (в этом случае происходит суммация процессов с противоположными знаками).
Длительные изменения МП, возникающие при синаптической активации, обнаружены в различных нервных клетках, в частности в МН спинного мозга.
Уже в одной из первых работ, проведенных на МН с помощью микроэлектродов, было показано, что при высокой частоте афферентной стимуляции из моносинаптических ВПСП может складываться плато деполяризации (Brock, Coombs, Eccles, 1952). В дальнейшем многие авторы наблюдали деполяризационное плато при высокочастотной синаптической активации, не проводя подробного анализа этого феномена (Eccles, 1953; R. Eccles, 1955; Araki a. Otani, 1959, и др.). Подбирая материал по характеристике синаптического электрогенеза, Г. Грундфест обратил специальное внимание на длительные изменения МП в результате суммации ПСП и впервые указал на связь между величиной деполяризационного плато и частотой афферентной стимуляции (Grundfest, 1957, 1959). Он подчеркнул, что аналогичные явления должны наблюдаться и при суммации ТПСП, не приводя соответствующих экспериментальных данных. Суммация полисинаптических ТПСП в МН спинного мозга с убыванием величины каждого последующего потенциала была показана А. И. Шаповаловым (1961а).
В настоящее время устойчивые сдвиги МП при высокочастотной синаптической активации изучены довольно подробно (Вартанян, 1962, 1964; Вартанян и Василевский, 1964б). При активации возбуждающих афферентных источников в МН возникает устойчивый деполяризационный сдвиг, который в определенном диапазоне растет пропорционально силе и частоте стимуляции (рис. 47). При высокочастотном раздражении тормозящих источников развивается гиперполяризационный сдвиг, подчиняющийся тем же закономерностям. Наибольшая величина устойчивых сдвигов возникает при некоторой частоте, определяемой длительностью переднего фронта ПСП, лежащего в основе устойчивого сдвига. Эта частота получила название предельной, поскольку дальнейшее увеличение частоты стимуляции приводит к уменьшению величины устойчивого сдвига (рис. 48). Чем короче передний фронт ПСП, лежащего в основе сдвига, тем выше предельная частота (Вартанян, 1964). Например, в случае моносинаптических ВПСП, обладающих коротким передним фронтом (1-1.5 мсек.), предельные частоты достигают 500-1000 в 1 сек. (рис. 47, II, и рис. 48, II).
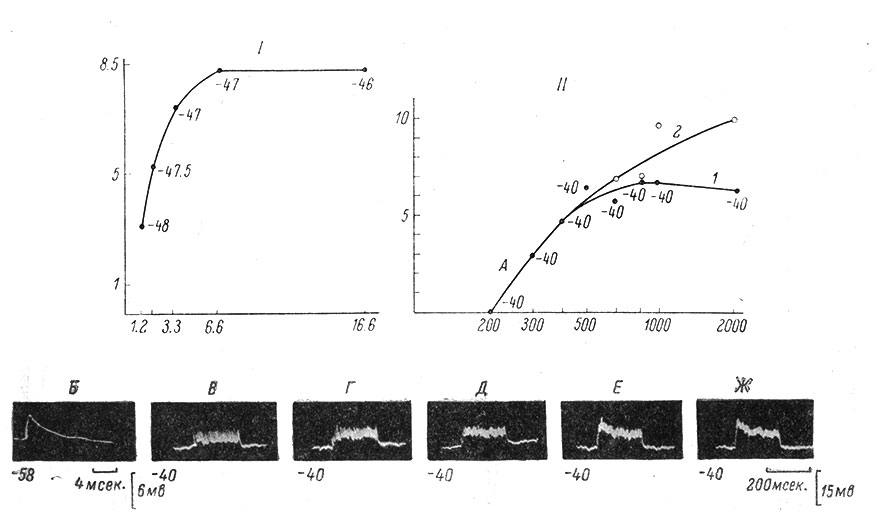
Рис. 47. Зависимость величины устойчивого деполяризационного сдвига мембранного потенциала от силы (I) и частоты (II) афферентной стимуляции. (Ориг.). I. По оси абсцисс - интенсивность стимуляции в порогах; раздражается n. semimembranosus с частотой 250 в сек. (длительность одиночного стимула 0.05 мсек.); по оси ординат - величина устойчивого деполяризационного сдвига, в мв; одиночный ВПСП, лежащий в основе устойчивого сдвига, имеет латентный период 6.4 мсек., длительность переднего фронта 4.6 мсек., амплитуду 2 мв. У точек на графике указана величина МП мотонейрона непосредственно перед нанесением высокочастотного раздражения длительностью 100 мсек. II - А - кривые зависимости величины устойчивого (1) и максимального (2) деполяризационного сдвига от частоты афферентной стимуляции. По оси абсцисс (логарифмическая шкала) - частота стимуляции; по оси ординат - величина деполяризационных сдвигов, в мв. Около точек на графике указана величина МП мотонейрона непосредственно перед нанесением ритмического высокочастотного раздражения длительностью 200 мсек.; здесь и во всех последующих записях раздражается n. biceps-semitendinosus (длительность одиночного стимула 0.05 мсек.; 1,5 порога моносинаптического ВПСП). Б - одиночный моносинаптический ВПСП, лежащий в основе устойчивого сдвига. Калибровка и отметка времени - справа и внизу. В начале записи - уровень МП мотонейрона (в мв), при котором была сделана запись. Длительность переднего фронта ВПСП - около 0.7 мсек. (сопоставить с максимумом устойчивого деполяризационного сдвига на А). В-Ж - часть отдельных записей деполяризационных сдвигов, на основании которых построен график на А. В - при частоте стимуляции 400 в 1 сек., Г - 500 в 1 сек., Д - 670 в 1 сек.; обратить внимание на появление максимального сдвига в начале записи, постепенно ниспадающего до устойчивого уровня; Е - 1000 в 1 сек.; Ж - 2000 в 1 сек. Калибровка и отметка времени общие для В-Ж. Внизу у каждой записи величина МП, в мв
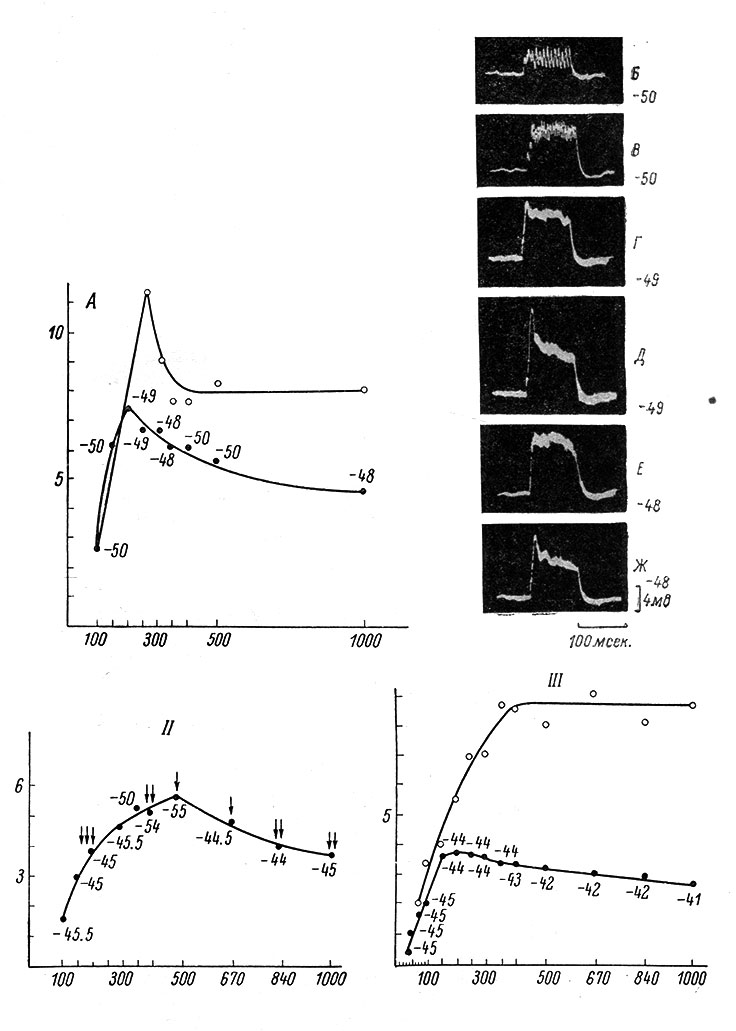
Рис. 48. Предельная частота афферентной стимуляции. (Ориг.). I. A - зависимость величины устойчивого (и максимального) деполяризационного сдвига, в основе которого лежит полисинаптический ВПСП (передний фронт = 4.6 мсек.), от частоты афферентной стимуляции. Обозначения осей для I, II, III те же, что на рис. 47, II, А, но по оси абсцисс - не логарифмический, а линейный масштаб. Черные кружки (то же для II, III) - устойчивый, белые - максимальный сдвиг. Около черных кружков показана величина МП мотонейрона, в мв (то же для II, III), непосредственно перед нанесением высокочастотных раздражений на n. semimembranosus (длительность одиночного стимула 0.05 мсек., интенсивность 1.6 порога). Наибольший устойчивый сдвиг имеет место при частоте 200 в 1 сек. Б-Ж - часть отдельных записей деполяризационных сдвигов, на основании которых построен график на А. Б - при частоте 100 в 1 сек., В - 150 в 1 сек., Г - 200 в 1 сек., Д - 250 в 1 сек., Е - 350 в 1 сек., Ж - 1000 в 1 сек. Калибровка и отметка времени для всех записей внизу справа. Справа у каждой записи - исходный МП мотонейрона, в мв. II - зависимость величины устойчивого деполяризационного сдвига, в основе которого лежит моносинаптический ВПСП (передний фронт = 1.5 мсек.), от частоты афферентной стимуляции. Стрелки - число ПД, возникающих в некоторых случаях при развитии деполяризационного сдвига. Раздражается n. gastrocnemius в течение 200 мсек. (сила одиночного стимула 1.5 порога, длительность 0.05 мсек.). Наибольший устойчивый сдвиг имеет место при частоте 500 в 1 сек. III - зависимость величины устойчивого (и максимального) деполяризационного сдвига, в основе которого лежит полисинаптический ВПСП (передний фронт = 6 мсек.), от частоты афферентной стимуляции. Раздражается n. biceps-semitendinosus в течение 200 мсек (сила одиночного стимула 25 порогов, длительность 0.05 мсек.). Наибольший устойчивый сдвиг имеет место в районе частот 150-250 в 1 сек
С помощью антидромного тестирования было показано, что при деполяризационных сдвигах возбудимость МН возрастает и он начинает воспроизводить более высокие ритмы стимуляции. На рис. 49 приведены индекс разряда МН при различных частотах антидромной стимуляции до (1) и во время деполяризационного сдвига (2), а также примеры записей, на основании которых построены графики. Видно, что индекс разряда МН заметно возрастает. При гиперполяризационных сдвигах возбудимость МН уменьшается и антидромный ПД может частично или полностью блокироваться (рис. 50).
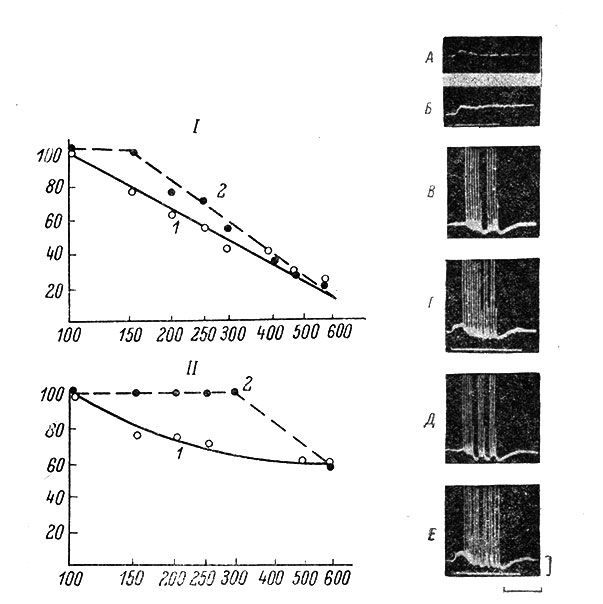
Рис. 49. Индекс разряда (ИР) мотонейрона при изолированной (2) антидромной ритмической стимуляции и в сочетании ее с устойчивым деполяризационным сдвигом (2). (Ориг.). I - индекс разряда, рассчитанный по полным антидромным ПД (состоящим из потенциалов начального сегмента НС- и сомато-дендритных СД-потенциалов). II - индекс разряда, рассчитанный с учетом всех ПД (как полных, так и состоящих только из НС-потенциалов). По оси абсцисс - частота антидромной стимуляции, в гц в логарифмическом масштабе; по оси ординат - индекс разряда. А - одиночный ВПСП, лежащий в основе устойчивого деполяризационного сдвига. Раздражается n. plantaris. Отметка времени на записи 20 мсек., МП = -60 мв. Б - устойчивый деполяризационный сдвиг при раздражении n. plantaris (4.4 порога) с частотой 80 в 1 сек. Устойчивый сдвиг в данном случае сохраняется и после прекращения стимуляции (последействие). МП = -58 мв. В - серия ПД в ответ на изолированную антидромную стимуляцию с частотой 200 в 1 сек. Видно полное выпадение нескольких ответов. В двух случаях ответ состоит только из НС-потенциала. МП = -56 мв. Г - то же, что и на Б, но на фоне устойчивого деполяризационного сдвига, вызванного раздражением n. plantaris с частотой 80 в 1 сек. (как на Б). Полного выпадения антидромных ответов нет. В трех случаях ответ состоит только из НС-потенциала. МП = -56 мв. Д - то же, что и на В, но частота антидромной стимуляции равна 250 в 1-сек. Видно полное выпадение нескольких ответов. В трех случаях ответ состоит только из НС-потенциала. МП = -53 мв. Е - то же, что и на Д, но на фоне устойчивого деполяризационного сдвига, вызванного раздражением n. plantaris частотой 80 в 1 сек. (как на Б). Полного выпадения антидромных ответов нет. В шести случаях ответ состоит только из НС-потенциала. МП = -53 мв. Внизу: калибровка для всех записей 20 мв; отметка времени - 70 мсек. для А, 160 мсек. для Б, 90 мсек. для В, Г, Д и Е. Время нанесения высокочастотного раздражения отмечено прямой белой линией. Все записи и графики относятся к одному и тому же МН
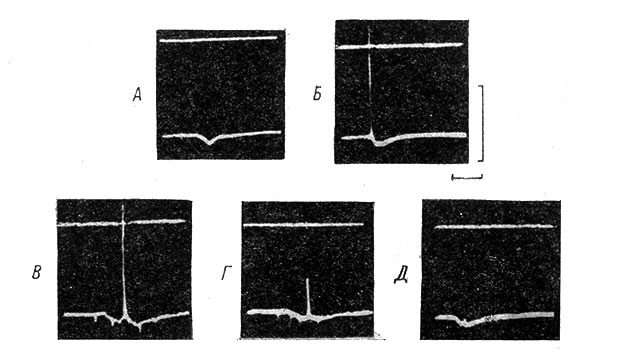
Рис. 50. Влияние гиперполяризационного сдвига на антидромный потенциал действия мотонейрона. (Ориг.). А - одиночный ТПСП, лежащий в основе устойчивого гиперполяризационного сдвига. Раздражается n. gastrocnemius. Б - одиночный антидромный ПД в ответ на раздражение переднего корешка L7. В - антидромное раздражение наносится на фоне гиперполяризационного сдвига, вызванного ритмической стимуляцией n. gastrocnemius с частотой 50 в 1 сек. Антидромный ответ не изменен. Г - то же, но при частоте афферентной стимуляции 75 в 1 сек. Сохраняется только НС-компонент антидромного ответа. Д - то же при частоте афферентной стимуляции 100 в 1 сек. Антидромный ответ блокируется полностью. Справа: калибровка для всех записей 50 мв, отметка времени 20 мсек. для А, 70 мсек. для Б, 40 мсек. для В, 30 мсек. для Г, 60 мсек. для Д. На всех записях вверху уровень 0 потенциала (отрицательность вниз). Все записи относятся к одному и тому же МН
Устойчивый деполяризационный сдвиг, повышая возбудимость МН, увеличивает вероятность импульсного ответа при ортодромной стимуляции. Небольшой ВПСП, который сам по себе не деполяризует нейрона до критического уровня, в условиях деполяризационного сдвига начинает генерировать ПД (рис. 51). Если амплитуда сдвига достаточно велика, то он может доводить МН до порогового уровня без дополнительных раздражений других афферентных источников (рис. 52). При этом в МН развивается импульсная активность, пропорциональная силе (рис. 52, А) и частоте (рис. 52, В) афферентной стимуляции.
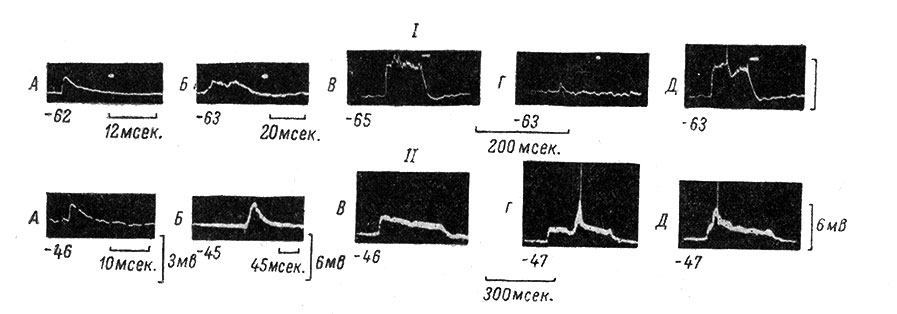
Рис. 51. Облегчающее влияние устойчивого деполяризационного сдвига на генерацию потенциала действия в мотонейроне. (Ориг.). I. A - моносинаптический ВПСП, возникающий в МН при одиночном раздражении n. gastrocnemius (сила 1.6 порога, длительность 0.05 мсек., отметка времени под записью; калибровка общая для всех записей). Внизу слева от всех записей - величина МП (в мв) в момент регистрации. Белая точка на всех записях - остановленный второй луч осциллографа, контролирующий уровень МП. Б - полисинаптический ВПСП в том же МН при одиночном раздражении n. peroneus (сила 5 порогов, длительность 0.05 мсек.). Отметка времени под записью. В - устойчивый деполяризационный сдвиг МП в том же МН при высоко частотном раздражении (670 в 1 сек.) n. gastrocnemius в течение 100 мсек. Отметка времени внизу, общая для В, Г и Д. Г - то же, что на Б, но при другой развертке. Д - то же, что на Б и Г, но на фоне устойчивого деполяризационного сдвига (как на В). В этом случае развитие одиночного полисинаптического ВПСП приводит к генерации ПД. II. А - моносинаптический ВПСП в другом МН, возникающий при одиночном раздражении n. biceps-semitendinosus (сила 2 порога, длительность 0.05 мсек.). Отметка времени на записи - 5 мсек. Калибровка справа. Внизу слева указан уровень МП, в мв (на всех других записях то же). Б - полисинаптический ВПСП в этом же МН в ответ на одиночное раздражение n. plantaris (сила 4 порога, длительность 0.05 мсек.). В - устойчивый деполяризационный сдвиг в том же МН при высокочастотном раздражении (670 в 1 сек.) n. biceps-semitendinosus в течение 300 мсек. Г, Д - одиночный полисинаптический ВПСП (при раздражении n. plantaris, как на Б) в разные периоды устойчивого сдвига (как на В). В обоих случаях полисинаптический ВПСП на фоне деполяризационного сдвига приводит к генерации ПД
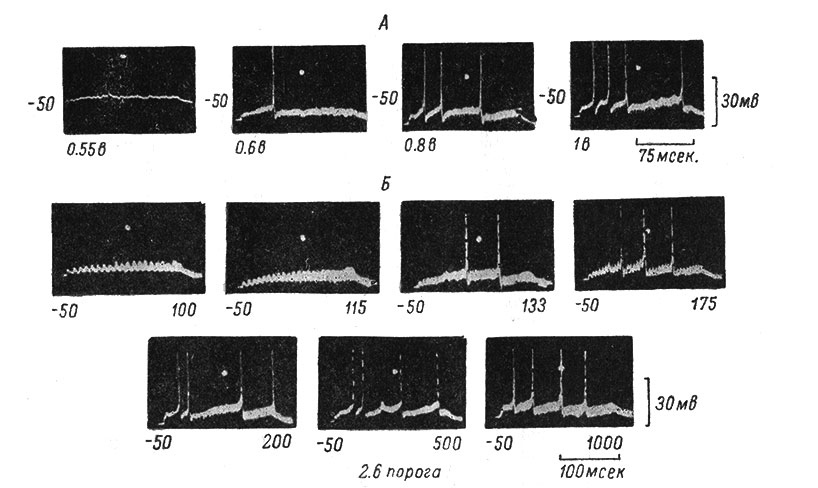
Рис. 52. Импульсная активность мотонейрона при различных величинах устойчивого деполяризационного сдвига мембранного потенциала. (Ориг.). А - влияние силы афферентного высокочастотного раздражения (670 в 1 сек.) на величину устойчивого деполяризационного сдвига и вызванную им импульсную активность. Слева направо даны записи устойчивого сдвига при усиливающейся интенсивности раздражения n. gastrocnemius (величина раздражения, в вольтах, дана под каждой записью). Около каждой записи слева дана исходная величина МП, в мв. Калибровка и отметка времени справа и внизу, общие для всех записей. Белая точка на записях - второй луч осциллографа, контролирующий величину МП. Видно, что с увеличением интенсивности раздражения число вызванных устойчивым сдвигом ПД возрастает. Б - влияние частоты афферентного моносинаптического раздражения (сила 0.8 в - 2.6 порога) на величину устойчивого деполяризационного сдвига и вызванную им импульсную активность (другой МН). Записи устойчивого сдвига при различной частоте (указана внизу справа от записей) раздражения n. gastrocnemius. Остальные обозначения те же, что на А. При частоте 200 в 1 сек. число пиков достигает уже максимума (4) и при дальнейшем увеличении частоты больше не растет
Гиперполяризационный сдвиг подавляет ортодромно вызванную импульсную активность МН, препятствуя достижению критического уровня деполяризации (рис. 53). В период гиперполяризационных устойчивых сдвигов возбудимость нейрона снижается. На рис. 54 показаны изменения МП при различных сочетаниях де- и гиперполяризационных сдвигов или устойчивых сдвигов и одиночных ПСП. Записи дают наглядное представление о том, как эти сдвиги могут менять возбудимость нейрона.
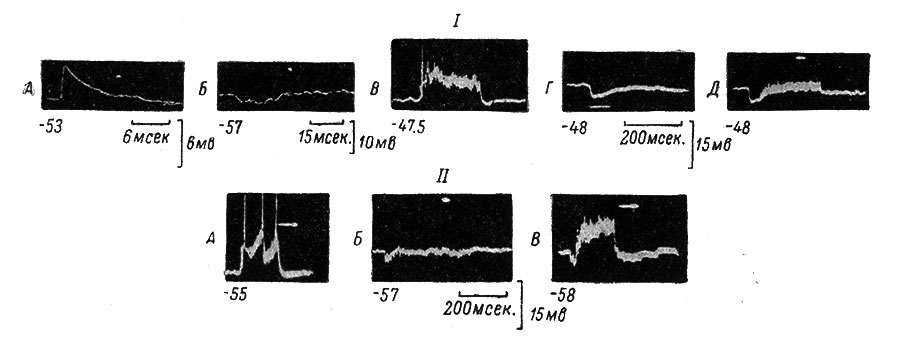
Рис. 53. Влияние гиперполяризационного устойчивого сдвига мембранного потенциала на синаптически вызванную импульсную активность мотонейрона. (Ориг.). I. А - моносинаптический ВПСП, возникающий в МН при одиночном раздражении n. biceps-semitendinosus (сила 4 порога, длительность 0.05 мсек.). Отметка времени под записью. Калибровка справа. Внизу слева от всех записей (А-Д и II) МП, в мв, в момент раздражения. Белая точка па записях - остановленный второй луч осциллографа, контролирующий уровень МН (и для II). Б - одиночный полисинаптический ТПСП в том же МН, возникающий при раздражении n. plantaris (сила 4 порога, длительность 0.05 мсек.). В - устойчивый деполяризационный сдвиг с одним ПД в начале высокочастотного раздражения (840 в 1 сек.) n. biceps-semitendinosus в течение 200 мсек. (тот же МН). Отметка времени и калибровка внизу; общие для В, Г и Д. Г - устойчивый гиперполяризационный сдвиг с выраженным последействием в том же МН при высокочастотном раздражении (240 в 1 сек.) n. plantaris (время раздражения отмечено на кадре непосредственно под записью). Д - сочетание двух высокочастотных раздражений: 840 в 1 сек. на n. biceps-semitendinosus и 240 в 1 сек. на n. plantaris (тот же МН). Видно, что гиперполяризационный сдвиг подавляет деполяризационный (начало записи) и препятствует развитию ПД. II. А - устойчивый деполяризационный сдвиг в том же МН с тремя ПД при высокочастотном раздражении (670 в 1 сек.) n. biceps-semitendinosus длительностью 200 мсек. Отметка времени и калибровка книзу, общие для А, Б и В. Б - гиперполяризационный сдвиг в том же МН, возникающий при высокочастотном раздражении (140 в 1 сек.) n. peroneus длительностью 500 мсек. В - сочетание обоих раздражений одновременно; деполяризационный сдвиг наложен на гиперполяризационный, в связи с чем величина первого по отношению к исходному уровню МП уменьшена и ПД отсутствуют (тот же МН)
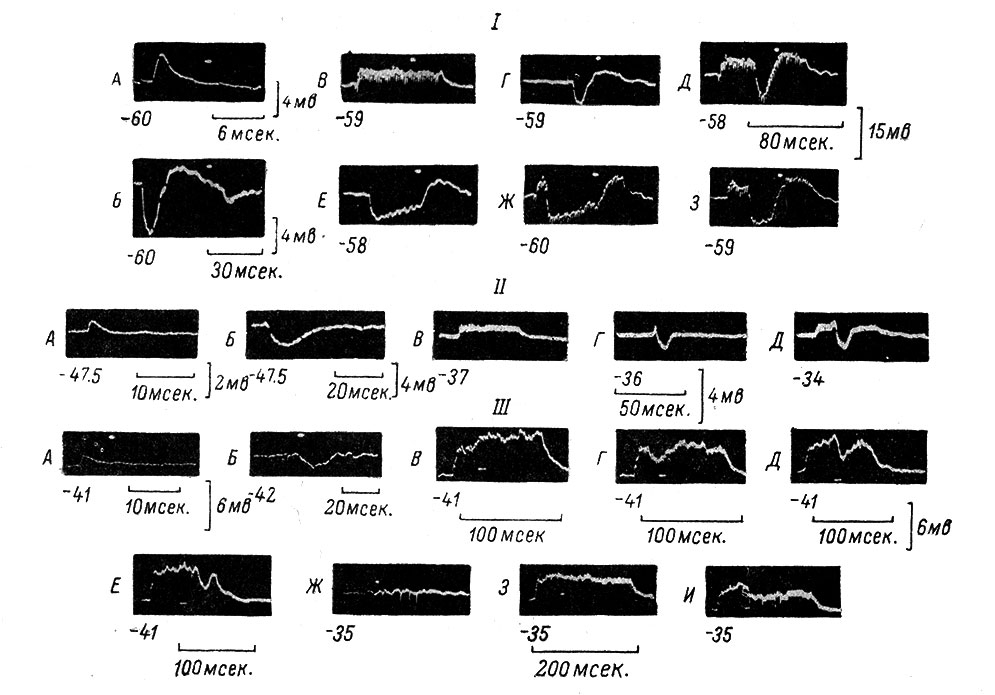
Рис. 54. Взаимодействие де- и гиперполяризационных устойчивых сдвигов между собой и с одиночными постсинаптическими потенциалами. (Ориг.). I. А - одиночный моносинаптический ВПСП, возникающий в МН при раздражении n. gastrocnemius (сила 2.3 порога, длительность 0.05 мсек.). Отметка времени и калибровка под записью. Внизу слева от этой и всех последующих записей - уровень МП, который контролируется вторым лучом осциллографа (белые точки па кадрах). Б - одиночный полисинаптический ТПСП с последующим переходом в полисинаптический ВПСП (преобладает тормозный эффект) в том же МН. Раздражается n. peroneus (сила 3.1 порога, длительность 0.05 мсек.). В - устойчивый деполяризационный сдвиг в том же МН, возникающий при высокочастотном раздражении (500 в 1 сек.) n. gastrocnemius в течение 100 мсек. Отметка времени и калибровка справа общие для В, Г, Д, Е, Ж и З. Г - то же, что и Б, но при другом усилении и замедленной развертке. Д - взаимодействие устойчивого деполяризационного сдвига (как на В) и одиночного комбинированного ПСП (как на Г). Тормозный эффект на фоне деполяризационного сдвига возрастает (ср. Д с Г). Е - гиперполяризационный сдвиг в том же МН, возникающий при высокочастотном раздражении (100 в 1 сек.) n. peroneus в течение 50 мсек. Ж - гиперполяризационный сдвиг (как на E) на фоне деполяризационного сдвига (как на В). Тормозный эффект на фоне деполяризационного сдвига возрастает (ср. Ж и Е). З - то же, что и Ж, но раздражение тормозящего афферентного источника (n. peroneus, 100 в 1 сек.) длится лишь 20 мсек. II. А - одиночный моносинаптический ВПСП в другом МН. Раздражается n. gastrocnemius (сила 1.55 порога, длительность 0.05 мсек.). Отметка времени и калибровка под записью. Слева внизу от этой и следующих записей - исходный уровень МП, в мв. Б - одиночный полисинаптический ТПСП в этом же МН. Раздражается n. peroneus (сила 7 порогов, длительность 0.05 мсек.). В - устойчивый деполяризационный сдвиг в том же МН, возникающий при высокочастотном раздражении (500 в 1 сек.) n. gastrocnemius, в течение 50 мсек. Отметка времени и калибровка общие для В, Г и Д, под записями. Г - то же, что и Б, но при другом усилении и замедленной развертке. Д - одиночный ТПСП (как на Г) на фоне деполяризационного сдвига (как на В) в том же МН. Тормозный эффект на фоне деполяризационного сдвига усиливается (ср. с Г). III. А - одиночный моносинаптический ВПСП в третьем МН. Раздражается n. biceps-semitendinosus (сила 1.25 порога, длительность 0.05 мсек.). Отметка времени и калибровка под записью. Б - одиночный полисинаптический ТПСП в этом же МН. Раздражается n. plantaris (сила 4 порога, длительность 0.05 мсек.). В - деполяризационный сдвиг в том же МН, возникающий при высокочастотном раздражении (670 в 1 сек.) n. biceps-semitendinosus в течение 100 мсек. Г - одиночный ТПСП (как на Б, но при другой развертке) на фоне деполяризационного сдвига (как на В). Д, Е - то же, что и Г, но при другой развертке. На Г-Е тормозное раздражение наносится с разной задержкой по отношению к началу деполяризационного сдвига. Ж - отсутствие видимого гиперполяризационного сдвига под влиянием высокочастотного раздражения (100 в 1 сек.) n. plantaris в течение 100 мсек. Отметка времени общая для Ж, З и И, внизу. З - то же, что и В, но при другой развертке. Длительность высокочастотного раздражения 200 мсек. И - проявление тормозного эффекта высокочастотного раздражения n. plantaris (как на Ж) на фоне деполяризационного сдвига (как на З)
Длительные синаптические влияния, не приводящие к возникновению импульсной активности, но вызывающие длительные сдвиги уровня МП, можно рассматривать как тонические. Они подготавливают нервную клетку к экстренным "фазическим" влияниям с других афферентных источников, создавая для последних благоприятные условия прохождения через данный нейрон к последующим. Если рассматривать нейрон как звено канала нервной связи, то можно сказать, что деполяризационный сдвиг оказывает на него оптимизирующее воздействие, повышая его пропускную способность. Он воспроизводит в частотном коде (в виде импульсов) даже небольшие деполяризационные процессы, так как устойчивый сдвиг приближает МП к критическому уровню. Уменьшается нестабильность импульсных разрядов, поскольку флуктуации ВПСП при этом перестают сказываться. Нейрон начинает пропускать более высокие пресинаптические ритмы, несмотря на то что при частой импульсации ВПСП уменьшаются по амплитуде (Curtis a. Eccles, 1960; Вартанян и Василевский, 1964, и др.).
Наоборот, гиперполяризационный сдвиг, возникающий при тормозящей стимуляции, может полностью или частично "запирать" работающий нейрон, прекращать его импульсацию и таким образом блокировать данное звено нервного канала связи.
Устойчивые сдвиги МП не следует рассматривать как чисто искусственное явление, не имеющее места в естественных условиях деятельности нейронов. Конечно, ритмическая активация не является типичной для деятельности центральных нейронов, однако высокие частоты импульсации с постоянно увеличивающимися интервалами между импульсами весьма распространены в различных отделах ц. н. с. Да и афферентная импульсация из рецепторных образований тоже зачастую характеризуется высокой частотой. Так, от аннуло-спиральных окончаний в спинной мозг нередко поступают импульсы с частотой 150-200 в 1 сек. (Matthews, 1933; Eldred, Granit a. Merton, 1953; Гранит, 1955, и др.). В отдельных случаях частоты достигают 500/сек.
Высокочастотная активация центральных нейронов в естественных условиях может также складываться в результате конвергенции относительно низкочастотной импульсации из нескольких афферентных источников. Фоновая активность ряда корковых нейронов достигает 150 импульсов в 1 сек. (Li a. Jasper, 1953). Высокочастотным разрядом обладают нейроны ретикулярной формации (Amassian a. De Vito, 1954; Лиманский, 1961, 1962). Возможно, именно эти высокочастотные разряды ретикулярных нейронов оказывают облегчающие и тормозящие влияния на нервные клетки других отделов ц. н. с. Такие влияния показаны в отношении двигательных и промежуточных нейронов спинного мозга, хотя при этом и не выявлено каких-либо изменений МП (Koizumi, Uschiyama a. Brooks, 1959). Однако стимуляция других надсегментарных образований (Sasaki, Namikawa a. Hashiramoto, 1960), в частности прецентральной извилины коры (Preston a. Whitlock, 1961), может вызывать длительные сдвиги поляризации МН. Имеются также данные о наличии длительных ПСП (десятки и даже свыше 100 мсек.) в пирамидных и непирамидных корковых нейронах при стимуляции срединных и вентролатеральных отделов таламуса (Purpura a. Shofer, 1964; Purpura, Shofer a. Musgrave, 1964). Наконец, в последнее время показано, что устойчивые сдвиги МП мотонейронов могут наблюдаться и в условиях адекватной стимуляции проприоцепторов мышц, причем эти сдвиги вызывают соответствующие изменения возбудимости МН по отношению к действию трансмембранного деполяризующего тока и ортодромной, синаптической стимуляции (Granit, Kellerth a. Williams, 1964a, 1964b).
Поэтому изучение устойчивых сдвигов МП на МП спинного мозга представляет интерес не только потому, что эти клетки являются удобной моделью для выяснения принципов регуляции возбудимости в центральном нейроне, но и в связи с тем, что такие сдвиги могут быть одним из важных фактически существующих механизмов ее регуляции.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'