II. Аккомодация
Надо сказать, что критический уровень деполяризации не является постоянной величиной. Длительная или медленно нарастающая деполяризация (Araki a. Otani, 1959; Frank a. Fuortes, 1960; Otani, 1960; Araki, Ito a. Oshima, 1961; Sasaki a. Otani, 1961, и др.), а также высокая частота импульсации (Eyzaguirre a. Kuffler, 1955; Kolmodin a. Skoglund, 1958) могут сдвигать критический уровень в сторону деполяризации, как бы "уводя" его от МП, и тем самым понижать возбудимость нейрона. Вопрос о соотношении уровня поляризации и возбудимости был поднят Н. В. Голиковым в 1933 г. В дальнейшем на основании анализа обширного экспериментального материала им было показано, что наиболее высокая возбудимость имеет место при некотором среднем ("нормальном") уровне поляризации нервной ткани. Как повышение, так и понижение нормальной поляризации приводит к снижению возбудимости и скорости проведения (Голиков, 1950).
В настоящее время снижение возбудимости во время длительной деполяризации связывают с возникающими при этом изменениями натриевой проводимости мембраны нейрона. Как уже указывалось выше (гл. 3, рис. 29), кривая зависимости специфической натриевой проводимости от уровня МП носит S-образный характер. Уровень МП определяет не только величину этой проводимости, но и степень ее инактивации. При медленной и длительной деполяризации степень инактивации увеличивается и соответственно уменьшается величина натриевой проводимости, которая может быть вызвана дополнительной быстрой деполяризацией. Кроме того, известную роль здесь может играть также повышение калиевой проводимости, возникающее под влиянием деполяризации. Оно увеличивает реполяризующие процессы и препятствует дополнительным деполяризующим влияниям.
Значительная деполяризация может полностью инактивировать специфическую натриевую проводимость, и тогда в нейроне возникает так называемый деполяризационный блок, или католическая депрессия, описанная на целом нервном стволе еще Б. Ф. Вериго в 1888 г.
Снижение возбудимости под влиянием длительной подпороговой или медленно нарастающей деполяризации получило название аккомодации.* Иногда используется также термин "адаптация", однако поскольку его чаще применяют для обозначения некоторых феноменов в деятельности рецепторных аппаратов, то лучше пользоваться первым.
* (Собственно говоря, этим понятием давно пользовались для обозначения соответствующих явлений в нервных стволах и мышцах. Лишь введение микроэлектродной техники позволило выявить их физиологический механизм.)
Аккомодационный процесс связан не только со сдвигом критического уровня, но и с частичным восстановлением МП при, действии длительной деполяризации. На рис. 45 (А, Б, В) приводятся данные из работы Араки, Ито и Ошима (Araki, Ito a. Oshima, 1961), которые наглядно это иллюстрируют (см. частичное восстановление МП и порога в ходе продолжающейся поляризации на В). Следует отметить, что аналогичные явления происходят также при применении гиперполяризующих токов (рис. 45, Б-В), что указывает на аккомодацию нейронов и к этим воздействиям.
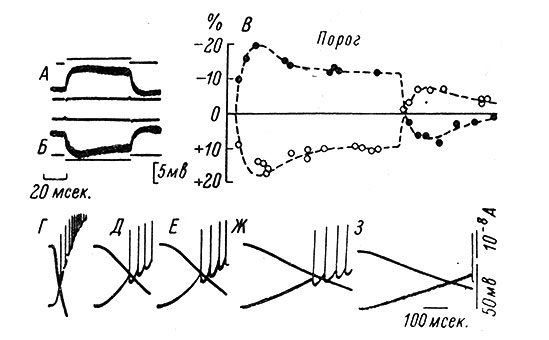
Рис. 45. Аккомодация нейронов. (Eccles, 1964). А, Б - деполяризационные и гиперполяризационные изменения мембранного потенциала мотонейрона кошки, вызванные прямоугольными толчками трансмембранного тока. Рядом с внутриклеточными записями даны соответствующие внеклеточные записи: под записью МП (на А) и над записью МП (на Б). В - изменения возбудимости другого МН по порогам коротких электрических раздражений во время и после пропускания длительных (93 мсек.) толчков де- (черные кружки) и гиперполяризующего (белые кружки) токов. По оси абсцисс - время; по оси ординат - уменьшение (вверх) и увеличение (вниз) порогов, в % к исходной величине. Изменения МП, вызванные этими токами, показаны в виде прерывистых линий и соответствуют изменениям МП на А и Б (Araki, Ito a. Oshima, 1961). Г-З - нижние записи представляют собой внутриклеточные потенциалы МН кошки при различной крутизне нарастания деполяризующих токов (верхние линии). Калибровка для внутриклеточных потенциалов и деполяризующих токов - справа. Отметка времени - внизу. (Sasaki a. Otani, 1961)
В последнее время аккомодационные процессы были довольно подробно изучены на МН спинного мозга кошки и крысы (Granit, Kernell a. Shortess, 1963). Используя длительные деполяризующие токи различной интенсивности, авторы обнаружили два типа аккомодации в МН. Одни МН характеризуются быстрой аккомодацией, давая при длительной деполяризации лишь несколько импульсных разрядов в самом начальном периоде действия деполяризующего тока. Эти МН рассматриваются авторами как фазические. Другие отвечают импульсной активностью на деполяризующий ток в течение всего периода его действия. Правда, после нескольких первых импульсов частота разрядов несколько уменьшается, но через некоторое время (около 1 сек.) она стабилизируется и может сохраняться в течение многих секунд (до 25). В целом частота следования импульсов после достижения устойчивых значений бывает в определенном диапазоне (до 64/сек. у крыс и до 37/сек. у кошек) пропорциональна силе деполяризующего тока. МН, характеризующиеся медленной и слабо выраженной аккомодацией, рассматриваются авторами как тонические.
Наиболее отчетливо закономерности аккомодационного процесса выявляются при использовании медленно нарастающих деполяризующих токов. Использование таких токов позволило, в частности, показать, что аксон и аксональный холмик МН кошек характеризуются быстрой аккомодацией, в то время как соматодендритная мембрана отличается более медленными аккомодационными процессами (Frank a. Fuortes, 1960; Sasaki a. Otani, 1961). Из рис. 45 (Г, Д, Е, Ж, З) видно, что, чем медленнее нарастает деполяризующий ток, тем больше латентный период первого импульса и тем большая деполяризация необходима для его появления. Таким образом, замедленная деполяризация особенно наглядно выявляет смешение критического уровня в деполяризационном направлении. Соотношение между степенью возрастания критического уровня деполяризации и латентным периодом первого импульса может служить мерой аккомодационного процесса (Araki a. Otani, 1959; Otani, 1960). Если при постепенном уменьшении крутизны деполяризующего тока критический уровень возрастает быстрее, а предел удлинения латентного периода первого импульса очень ограничен (дальше генерация ПД делается невозможной), то аккомодация происходит быстро. Если же в этих условиях критический уровень увеличивается медленно, а латентный период первого импульса имеет возможность существенно нарастать, то аккомодационный процесс носит медленный характер (рис. 46).
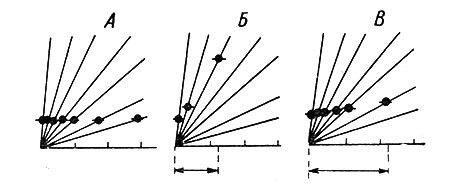
Рис. 46. Скорость возрастания критического уровня деполяризации при быстрой (Б) и медленной (В) аккомодации. (Ориг.). А - аккомодация отсутствует. Расходящиеся из одной точки линии схематически изображают разную крутизну нарастания деполяризации. Черные точки - критический уровень деполяризации. Стрелки внизу - латентный период первого импульса при минимальной крутизне деполяризации, еще способной вызвать ПД, в случаях быстрой и медленной аккомодации
Таким образом, в каждый данный момент возбудимость нейрона определяется соотношением величины МП и критического уровня деполяризации. При относительно коротких и быстро развивающихся изменениях МП аккомодационные процессы не оказывают существенного влияния на возбудимость нейрона, и она практически зависит от уровня МП. Конечно, при значительных деполяризациях эти соотношения нарушаются, потому что при развитии деполяризационного блока нейрон не способен генерировать импульсы, и в таких условиях не существует ни критического уровня деполяризации, ни возбудимости.
Чем же определяется уровень МП в нейроне?
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'