Глава 4. Синаптическая передача нервных процессов (Г. А. Вартанян)
Импульсная активность, возникающая в нейроне в результате развития сверхпороговой деполяризации, вызывает в конечном итоге секрецию медиатора в аксональных окончаниях этого нейрона. Правда, в последнее время обнаружен ряд межнейрональных контактов, в которых имеет место чисто электрическая передача нервных процессов (Hagiwara a. Bullock, 1957; Furshpan a. Potter, 1959; Martin a. Pillar, 1963, и др.). Однако в подавляющем большинстве рецепторно-нейрональных, межнейрональных и нейронально-эффекторных соединений самых различных животных передача осуществляется с помощью химических посредников, или медиаторов. По-видимому, в ходе эволюции центральные нейроны развились из первичных секреторных клеток (Грундфест, 1961а), и нервная деятельность сохранила химическую основу на всех этапах эволюции, включая человека (Раштон, 1961).
Основные механизмы протекания нервных процессов в постсинаптическом звене синаптического контакта были рассмотрены в предыдущей главе. Поэтому здесь необходимо рассмотреть главным образом процессы, развивающиеся в пресинаптическом звене синапса, и явления, связанные с секрецией медиатора в синаптическую щель.
Электронномикроскопические исследования последнего десятилетия показали, что в районе контакта между пресинаптическим окончанием и субсинаптической мембраной, по обе стороны синаптической щели имеются утолщенные и уплотненные участки обеих мембран (пре- и субсинаптической). В утолщенных участках пресинаптических окончаний сосредоточено большое количество своеобразных органоидов, получивших название синаптических пузырьков, или везикул. Как уже указывалось в главе 2, в настоящее время эти везикулы рассматриваются в качестве квантовых скоплений медиатора. Приход нервного импульса в пресинаптическое окончание сопровождается синхронным выбросом в синаптическую щель содержимого большого числа пузырьков, расположенных в непосредственной близости от нее. Утолщенный участок субсинаптической мембраны представляет собой рецепторную зону, снабженную специфическими рецепторными молекулами, с которыми взаимодействуют выброшенные в синаптическую щель молекулы передатчика. В результате этого взаимодействия структура субсинаптической мембраны временно изменяется. В ней возникают "поры" определенной величины. Они достаточно велики в возбуждающих синапсах и пропускают все виды низкомолекулярных ионов. Таким образом, сопротивление субсинаптической мембраны оказывается резко сниженным, благодаря чему постсинаптическая мембрана как бы шунтируется. Возникает ионный ток, направление которого было показано ранее на рис. 23, и в нейроне развивается ВПСП (центральное возбуждение). В тормозящих синапсах возникают "поры" меньшего диаметра, около 2.5 Å, через которые могут проходить только некоторые ионы. При этом в постсинаптической мембране возникают токи обратного направления - гиперполяризующие, которые лежат в основе ТПСП (центральное торможение).* Действие медиатора кратковременно, поскольку он вскоре разрушается специфическим ферментом, функционирующим в районе синаптического контакта.
* (См. подробности в предыдущей главе.)
Какие же экспериментальные данные послужили основанием для этих схематически изложенных представлений?
Механизм синаптической передачи наиболее подробно изучен в нервно-мышечном соединении. Однако есть достаточно оснований считать, что в центральных синаптических аппаратах секреторно-передаточные процессы имеют сходные закономерности (Мс Lennan, 1963; Eccles, 1964; Шаповалов, 1964).
В 1952 г. Фэтт и Катц (Fatt a. Katz, 1952) записали так называемые миниатюрные потенциалы концевой пластинки (мин. ПКП) в районе нервно-мышечного соединения. Эти потенциалы по различным характеристикам оказались очень похожими на обычные ПКП. Однако по амплитуде они приблизительно в сто раз меньше последних и наблюдаются в покое, когда в двигательном нерве импульсация отсутствует (рис. 38). Исследователи прежде всего обратили внимание на тот факт, что мин. ПКП, так же как и ПКП, вызванные нервными импульсами, увеличиваются и удлиняются под действием антихолинэстеразных веществ и блокируются курареподобными препаратами (Fatt a. Katz, 1952; Del Castillo a. Katz, 1956b, и др.). Далее оказалось, что мин. ПКП возникают по случайному закону; при деполяризации пресинаптических окончаний частота их появления значительно возрастает, а при гиперполяризации падает (Del Castillo a. Katz, 1954b; Liley, 1956b) (рис. 39, A, Б, В). Лили (Liley, 1956b), в частности, показал, что избыток ионов К+ в окружающей среде (и возникающая в связи с этим деполяризация пресинаптических окончаний) приводит к значительному увеличению частоты мин. ПКП (рис. 39, Г).
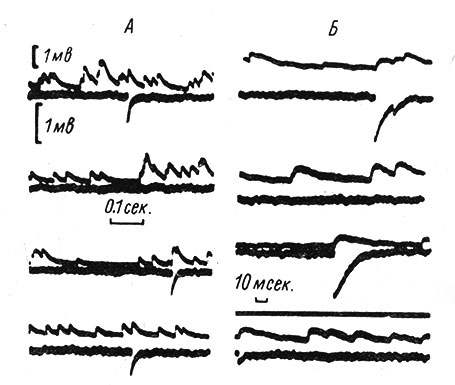
Рис. 38. Миниатюрные потенциалы концевой пластинки (ПКП). (Del Castillo a. Katz, 1956а). А, Б - одновременная внутри- и внеклеточная запись спонтанных мин. ПКП мышечного волокна лягушки. Верхняя кривая в каждой записи зарегистрирована внутриклеточным электродом. Для всех записей отклонение вверх регистрирует положительность. Б - записи сделаны при большей скорости развертки луча. Калибровки: верхняя - для внутриклеточной записи, нижняя - для внеклеточной
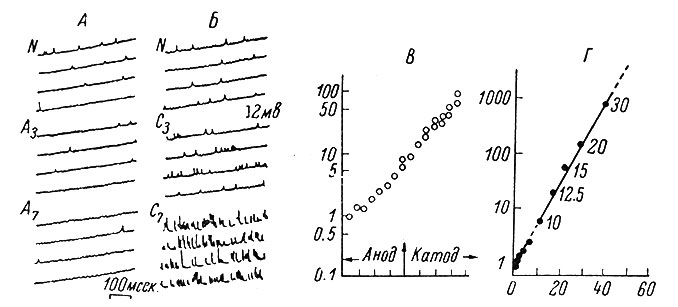
Рис. 39. Зависимость частоты мин. ПКП от уровня поляризации пресинаптических окончаний. (Liley, 1956b). А, Б - частота мин. ПКП (внутриклеточная запись от одиночного мышечного волокна диафрагмы крысы) до (N) и после гипер- (А3, А7) или деполяризующих (С3, С7) токов, проходящих через нервное волокно и электротонически меняющих уровень поляризации пресинаптических окончаний. Цифры у символов - 3 и 7 - указывают на относительную интенсивность гипер- (А) и деполяризующего (С) токов. В - зависимость частоты мин. ПКП (ось ординат) от интенсивности поляризующего тока в относительных единицах (ось абсцисс). Г - зависимость между частотой мин. ПКП (ось ординат) и высчитанными значениями деполяризации (в мв) пресинаптических окончаний, возникшей под влиянием повышенной концентрации калия в окружающей среде (величина концентрации калия указана у соответствующих точек в мМ). График построен Б. Катцем (Katz, 1962)
Особенно обращает на себя внимание тот факт, что под влиянием различных воздействий изменяется только частота возникновения мин. ПКП, но не их величина (амплитуда). Постоянной амплитуде мин. ПКП соответствует одинаковая величина синаптических везикул, расположенных в непосредственной близости от синаптической щели, в так называемой оперативной зоне.* Если допустить, что ацетилхолин находится в везикулах в изотонической концентрации, что кажется наиболее вероятным, ото можно подсчитать, что в одном пузырьке диаметром 500 Å должно содержаться около 6000 молекул ацетилхолина (Eccles, 1964), что соответствует экспериментальным определениям количества ацетилхолина, вызывающего один мин. ПКП (Del Castillo a. Katz, 1954а, 1956b; Katz, 1958). Кроме того, распределение синаптических пузырьков в пресинаптических окончаниях соответствует множественности точек возникновения ПКП в районе синаптического контакта, что было показано Кастильо и Катцем с помощью внутриклеточного отведения (Del Castillo a. Katz, 1956a). В связи с этим было сделано заключение, что мин. ПКП связаны с выделением в синаптическую щель более или менее одинаковых порций, квант, ацетилхолина - точно установленного медиатора нервно-мышечной передачи (см. подробно: Katz, 1962).
* (Термин "оперативная зона" представляется здесь более соответствующим сути дела, чем принятый в англо-американской литературе термин "стратегическая зона" (strategic zone).)
Наличие везикул в пресинаптических терминалях обнаружено во многих межнейрональных синаптических контактах (De Robertis, 1958, 1959; Palay, 1958a; Whittaker a. Gray, 1962). Этим морфологическим находкам соответствуют и нейрофизиологические данные о мин. ПСП в различных нейронах. Правда, при исследовании мин. ВПСП в центральных нейронах в большинстве случаев невозможно исключить асинхронную синаптическую "бомбардировку", которая может "симулировать" мин. ВПСП. Однако в некоторых случаях наличие мин. ПСП в центральных нейронах можно считать доказанным, например, в мотонейронах изолированного спинного мозга лягушки при подавлении рефлекторной деятельности мозга высокой концентрацией ионов Mg++ в омывающей среде (Katz a. Miledi, 1963), а также в клетках изолированного симпатического ганглия, где случайная синаптическая "бомбардировка" полностью исключается (Nishia. Koketsu, 1960; Blackman. Ginsborg a. Ray, 1962). В то же время следует отметить, что отсутствие мин. ПСП в центральных нейронах не может говорить против химической передачи нервных процессов в синапсах. По-видимому, в этих случаях пресинаптические окончания не имеют базальной секреции, и выделение медиатора происходит только при определенном уровне деполяризации пресинаптического окончания (Eccles, 1964).
При поступлении нервного импульса в пресинаптические окончания происходит интенсивное выделение медиатора в синаптическую щель. Почти совершенно синхронно освобождается большое число квант медиатора, о чем можно судить на основании сравнения мин. ПКП с ПКП, вызванным нервным импульсом (Katz, 1958, 1962). То, что при деполяризации пресинаптических окончаний нервным импульсом выделение медиатора в синаптическую щель носит квантовый характер, почти не вызывает сомнений в связи со следующими экспериментальными данными.
Известно, что для возникновения нормального ПКП необходимо определенное количество ионов Са++ в межклеточной жидкости. Недостаток кальция снижает и может частично или полностью блокировать ПКП (Del Castillo a. Katz, 1954a; Liley, 1956b). Аналогичное влияние на ПКП оказывает избыток магния (Del Castillo a. Engback, 1954; Jenkinson, 1957). Ионы Mg++ рассматриваются как конкурентный агент по отношению к Са++ (Katz, 1962). Если уменьшить внеклеточную концентрацию кальция или увеличить концентрацию магния, то ПКП резко уменьшаются в размерах и начинают ступенчато флуктуировать от величин, близких к амплитуде мин. ПКП, до некоторых кратных ей величин (Del Castillo a. Katz, 1954b; Boyd a. Martin, 1956a; Liley, 1956a). По-видимому, эти флуктуации связаны с числом выделяющихся квант ацетилхолина (рис. 40). Использование статистических методов анализа (теоремы Пуассона) выявило аналогичные закономерности и в других типах синапсов, например, в нервно-мышечном соединении ракообразных (Dudel a. Kuffler, 1961a) и в синаптических переключениях симпатического ганглия лягушки (Blackmann, Ginsborg a. Ray, 1962). Расчеты с использованием различных методов показывают, что во время развития ПКП в синаптическую щель выделяется от 100 до 300 квант ацетилхолина (Del Castillo a. Katz, 1954a; Martin, 1955: Boyd a. Martin, 1956b; Liley, 1956a; Burke, 1957).
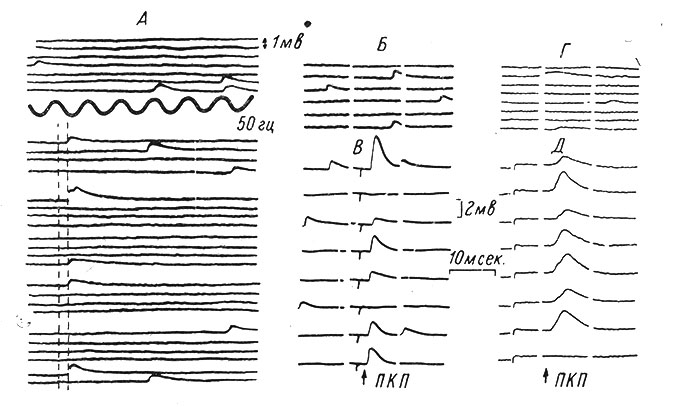
Рис. 40. Квантовые составляющие ПКП. А - внутриклеточная запись с нервно-мышечного соединения лягушки, показывающая несколько спонтанных мин. ПКП (верхние записи) и ответы на одиночные нервные импульсы при погружении препарата в раствор с недостаточным количеством кальция и избытком магния (нижние записи). Латентный период ответов отмечен двумя вертикальными прерывистыми линиями. Во многих случаях стимуляции ответы отсутствовали, и было только 5 ответов на 24 импульса (Del Castillo a. Katz, 1954а). Б-Д - внутриклеточные записи от мышечных волокон диафрагмы крысы; Б - контрольная запись спонтанных мин. ПКП для В; Г - контрольная запись для Д. В и Д - одиночные ПКП в ответ на одиночное раздражение нерва. Окружающий раствор содержит пониженное количество кальция (1 мМ) и избыток магния (6 мМ) (Liley, 1956b)
Вопрос о том, доходит ли нервный импульс (ПД) до аксональных терминалей или блокируется в местах разделения аксона на конечные ветви, только электротонически деполяризуя окончания, остается пока нерешенным. Ряд авторов полагает, что импульс блокируется на некотором расстоянии от пресинаптических окончаний. В качестве одного из доказательств пресинаптического блокирования сторонники этой точки зрения используют тот факт, что при высоких частотах стимуляции ПКП начинают воспроизводиться в трансформированном ритме (Шаповалов, 1964). Однако в одной из недавних работ, проведенной на нервно-мышечном соединении диафрагмы крысы, Хаббард и Шмидт (Hubbard a. Schmidt, 1963) с помощью внеклеточной регистрации активности пресинаптических окончаний показали, что ПД в большинстве случаев достигает конечных участков аксона. В то же время на других объектах: нервно-мышечном соединении ракообразных (Dudel a. Kuffler, 1961a), синаптическом переключении цилиарного ганглия цыпленка (Martin a. Pillar, 1963), - эти данные не подтверждаются. По-видимому, в различных синаптических аппаратах имеет место различная степень проникновения импульса в пресинаптические окончания. И в тех случаях, когда он не достигает самых конечных ветвей аксона, электротоническое деполяризующее влияние оказывается вполне достаточным для выброса медиатора в синаптическую щель (Eccles, 1964).
Механизм влияния деполяризационного процесса на скорость и синхронность выделения медиатора тоже остается пока еще неизвестным. На этот счет существует несколько гипотез. Дель Кастильо и Катц, наиболее компетентные исследователи в этой области, предполагают, что синаптические пузырьки находятся в постоянном тепловом движении. На них имеются рецепторные участки, стереометрически пригнанные к аналогичным рецепторным зонам внутренней поверхности пресинаптической мембраны. Взаимодействие, "замыкание", этих участков между собой является предварительным условием выделения содержимого пузырьков в синаптическую щель. Деполяризация значительно увеличивает количество рецепторных зон внутренней поверхности мембраны. В результате число взаимодействий между синаптическими везикулами и рецепторными зонами мембраны резко возрастает, и большое число квант ацетилхолина синхронно выделяется в синаптическую щель. Ионы Са++ являются необходимым кофактором для развития этого процесса (Del Castillo a. Katz, 1956b; Katz, 1958, 1962). Несомненно, что эти предположения весьма гипотетичны, однако они объясняют многие из полученных к настоящему времени экспериментальных данных.
Так или иначе амплитуда ПСП связана непосредственно с величиной пресинаптического ПД. В опытах на гигантском синапсе звездчатого ганглия кальмара было показано, что увеличение пресинаптического ПД (вызванное гиперполяризацией пресинаптического волокна) сопровождается увеличением ВПСП, а уменьшение импульса в пресинаптическом волокне (при его частичной деполяризации) вызывает снижение ВПСП (Hagiwara a. Tasaki, 1958; Takeuchi a. Takeuchi, 1962). Связь между величиной пресинаптического ПД и амплитудой ВПСП носит характер показательной функции (рис. 41).
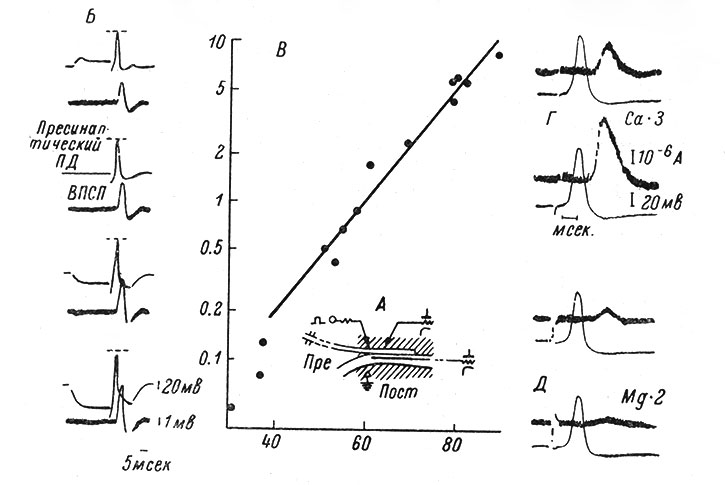
Рис. 41. Зависимость амплитуды ВПСП от величины пресинаптического нервного импульса (ПД). А - схема опытов Хагивара и Тасаки. Изображен гигантский синапс звездчатого ганглия кальмара с двумя электродами (поляризующим и регистрирующим) в гигантском пресинаптическом волокне и одним электродом в гигантском постсинаптическом волокне (Hagiwara a. Tasaki, 1958). Б-Д - соотношение между величиной пресинаптического ПД и амплитудой ВПСП (Takeuchi a. Takeuchi, 1962). Б - примеры внутриклеточных записей на гигантском синапсе звездчатого ганглия кальмара. Расположение электродов показано на А. Пресинаптический ПД (верхняя кривая) и генерируемый им ВПСП (нижняя кривая) показаны в конце развертки. В начале развертки 1-й записи пресинаптическое волокно деполяризуется прямоугольным толчком выходящего тока. В двух последних записях пресинаптическое волокно гиперполяризуется прямоугольными толчками входящего тока различной интенсивности. Прерывистые линии - абсолютная высота контрольного пресинаптического ПД. В - график зависимости амплитуды ВПСП от величины пресинаптического ПД на основании данных, частично представленных на Б. По оси абсцисс - величина ПД, в мв; по оси ординат - величина ВПСП в мв, в логарифмическом масштабе. Г - пресинаптические ПД (нижние кривые) и токи, вызывающие развитие ВПСП (верхние кривые), для демонстрации влияния повышенной в 3 раза концентрации ионов Са++ на амплитуду ВПСП; величина ПД при этом не изменяется. Д - подавляющее влияние удвоенной концентрации ионов Mg++ на токи, вызывающие ВПСП. Г, Д - использована методика фиксации мембранного потенциала
Сходные результаты были получены также на нервно-мышечном препарате диафрагмы крысы (Hubbard a. Willis, 1962a, 1962b, 1962с). Однако в этих опытах обнаружилось, что гиперполяризация пресинаптических окончаний вызывает гораздо большее увеличение ПКП, чем это можно было ожидать в результате увеличения амплитуды пресинаптического ПД. В то же время деполяризация окончаний вызывает снижение ПКП, пропорциональное интенсивности деполяризующего тока. По-видимому, гиперполяризация пресинаптических окончаний сопровождается процессом мобилизации передатчика, о чем речь будет идти в главе 5.
Таким образом, пресинаптический разряд вызывает интенсивное выделение медиатора, который, диффундируя в синаптическую щель, достигает рецептивных молекул субсинаптической мембраны и взаимодействует с ними. Именно с диффузией медиатора в настоящее время в основном связывают синаптическую задержку - характернейшую особенность синаптического проведения.
Экклс (Eccles, 1964) отмечает следующие составные части синаптической задержки: время нарастания переднего фронта пресинаптического ПД до некоторой величины, по достижении которой начинает выделяться существенное количество медиатора; время диффузии выделенного пресинаптическим окончанием медиатора к рецептивным зонам субсинаптической мембраны (по-видимому, это самая значительная составляющая синаптической задержки, достигающая 0.1 мсек.); и, наконец, время, необходимое для взаимодействия выделенных молекул передатчика с рецептивными молекулами субсинаптической мембраны и возникновения в ней повышенной ионной проницаемости (эта составляющая тоже может длиться довольно долго - около 0.1 мсек.).
Общая длительность синаптической задержки в нервно-мышечном соединении млекопитающих составляет 0.22 мсек. (Hubbard a. Schmidt, 1963), а в центральных синапсах, по данным различных авторов, колеблется в пределах 0.2-0.5 мсек. - главным образом в зависимости от используемых методов ее определения (Eccles, 1964). Точное определение длительности каждой составляющей синаптической задержки пока остается невозможным.
Согласно современным представлениям, молекулы медиатора, взаимодействуя с рецепторными молекулами субсинаптической мембраны, вызывают изменения ее структуры, переводя ее в "ситоподобное состояние". Конечно, это всего навсего крайне упрощенная схема происходящих в субсинаптической мембране процессов. Возникающие при этом молекулярные изменения мембраны все еще остаются совершенно неизвестными. Поэтому вряд ли целесообразно разбирать здесь многочисленные, но малодоказанные гипотезы о механизмах взаимодействия передатчика с субсинаптической мембраной. Важно, что под влиянием молекул передатчика в мембране постсинаптического нейрона действительно возникают выраженные изменения проницаемости, приводящие к развитию в нем возбуждения или торможения; в результате взаимодействия последних в нейроне возникает сложный рисунок импульсной активности.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'