Глава 5. Пластичность синаптической передачи (Г. А. Вартанян и Н. Н. Василевский)
Функциональные возможности синапсов не являются строго постоянными. При некоторых условиях их эффективность может изменяться, и вызванные пресинаптическим разрядом ПСП могут возрастать или уменьшаться.
В 1949 г. Дэвид Ллойд (Lloyd) показал, что после высокочастотного (500-600/сек.) раздражения волокон группы IA икроножного нерва рефлекторный ответ на одиночное тестирующее раздражение, записываемый с переднего корешка, значительно возрастает. Увеличенный рефлекторный ответ может сохраняться в течение нескольких минут после окончания высокочастотной стимуляции, постепенно возвращаясь к исходной величине (рис 42). Описанное явление получило название постактивационного усиления, или посттетанической потенциации (ПТП).
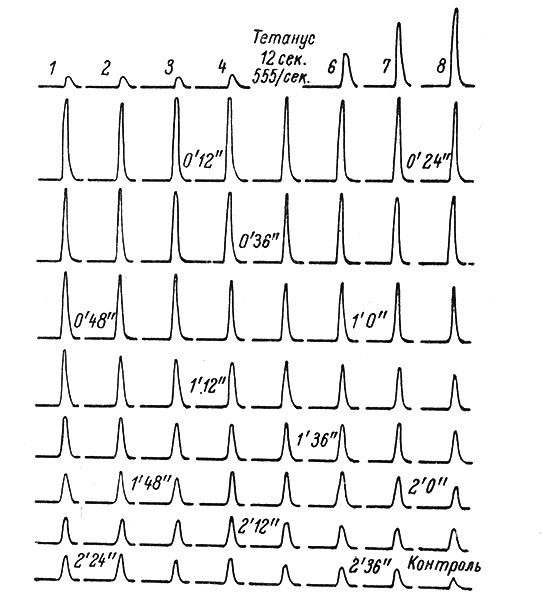
Рис. 42. Посттетаническая потенциация. (Lloyd, 1949). Моносинаптический рефлекторный ответ на одиночное раздражение записывается с переднего корешка S1. Раздражаются афферентные волокна I группы икроножного нерва. Записи 1-4 сделаны последовательное интервалом 2.4 сек. Между 4-й и 6-й записью проведена высокочастотная стимуляция нерва. Последующие записи одиночных ответов производились последовательно с интервалом 2.4 сек
Уже прохождение одного или нескольких импульсов через синапсы сопровождается кратковременной потенциацией, которая иногда оказывается лишь относительной, поскольку в пресинаптическом аппарате одновременно развивается также и депрессия (Curfis a. Eccles, 1960, и мн. др.). При короткой ритмической стимуляции (несколько сотен мсек.) постактивационный эффект зависит в основном от частоты раздражения. Обычно это потенциация при высоких частотах (порядка нескольких сот в секунду) и депрессия или только относительная потенциация (на фоне депрессии) при более низких частотах (ниже 100-1500 в 1 сек.). При длительной ритмической стимуляции (несколько секунд) потенциация развивается медленней, но оказывается более интенсивной и резко удлиняется. Если при кратковременной стимуляции потенциация длится в течение нескольких секунд, то при длительной она продолжается минутами и может развиваться при более низких частотах (100 в 1 сек.).
Природа и механизм обоих постактивационных эффектов - потенциации и депрессии - изучены еще крайне недостаточно, несмотря на большое число проведенных в этом направлении работ (Eccles, 1964).
Экспериментальный анализ этого феномена показал, что ПТП развивается в пресинаптических окончаниях, а не в теле нейрона. Ллойд связывал повышенную активность пресинаптических окончаний с их гиперполяризацией, которая закономерно возникает при высокочастотной активации (Lloyd, 1949; Eccles a. Rail, 1951; Wall a. Jhonson, 1958, и др.). Повышение МП пресинаптических волокон должно сопровождаться увеличением амплитуды проходящего по ним импульса, что может обусловливать большую эффективность соответствующего синаптического поля.
Действительно, последующие микроэлектродные исследования во многом подтвердили гипотезу Ллойда. Как уже указывалось в главе 4, исследования на гигантском синапсе кальмара показали, что гиперполяризация пресинаптического волокна вызывает увеличение пресинаптического ПД, что в свою очередь сопровождается появлением увеличенного ВПСП в постсинаптическом волокне. Поскольку связь между амплитудой пресинаптического ПД и величиной ВПСП имеет характер показательной функции, некоторые исследователи стали рассматривать эту зависимость как причину ПТП (Liley, 1956b). Сходные результаты были получены и на МН спинного мозга с помощью поляризации пресинаптических окончаний первого нейрона постоянным током (рис. 36). Деполяризация пресинаптических окончаний сопровождалась уменьшением ВПСП, а гиперполяризация - увеличением (Eccles, Kostyuk, Schmidt, 1962c).
Однако исследования последних лет позволяют считать, что увеличение амплитуды пресинаптического импульса не может быть определяющим фактором развития ПТП. По-видимому, доминирующим механизмом этого феномена является мобилизация медиатора в пресинаптических окончаниях под влиянием их гиперполяризации (Hubbard a. Willis, 1962a; Hubbard a. Schmidt, 1962, 1963, и др.). Возможно, что эта мобилизация имеет электрофоретический механизм, когда ионы медиатора перемещаются по направлению к "оперативной зоне" пресинаптического окончания под влиянием гиперполяризующего тока (Eccles, 1964). Механизм мобилизации медиатора, вероятно, не связан с механизмом выброса синаптических пузырьков, потому что гиперполяризация пресинаптических окончаний сопровождается не увеличением, а уменьшением частоты миниатюрных ПКП (Del Castillo a. Katz, 1954b; Liley 1956b, и др.). При высокой частоте следования пресинаптических импульсов мобилизационный процесс резко возрастает. Поэтому при прекращении высокочастотной стимуляции (сотни импульсов в секунду), когда происходит внезапная остановка интенсивного выделения медиатора, возникают условия для его накопления в самых конечных участках пресинаптического окончания и вновь приходящий через некоторое время импульс вызывает повышенный выход медиатора в синаптическую щель, в результате чего ВПСП резко возрастает (Curtis a. Eccles, 1960; Eccles, 1964). Однако в случае длительной ритмической стимуляции известную роль может играть также возникающая следовая гиперполяризация механизм действия которой описан выше.
Так или иначе в настоящее время имеется достаточно оснований, чтобы говорить о влиянии гиперполяризующих токов на обменные процессы в нервном волокне. Такая точка зрения находит подтверждение в последних электронномикроскопических исследованиях, в которых образование синаптических везикул связывается с деятельностью митохондрий концевого волокна (Давыдова, Дьячкова и Смирнов, 1965). С другой стороны, имеются данные, указывающие на то, что формирование синаптических пузырьков начинается в области перикариона, откуда они транспортируются по аксону в синаптические окончания (Van Breemen, Anderson a. Reger, 1958). Подобный механизм образования синаптических везикул весьма вероятен, так как имеются данные о центробежном перемещении в аксоплазме не только ацетилхолина, но и холинацетилазы, холинестеразы и ряда других метаболитов и ферментов (Feldberg, 1945; Sawjer, 1946; Banister a. Serase, 1950; Hebb a. Waites, 1956; Кибяков, 1964, и др.). По существу это уже трофические механизмы нервного волокна, которые тоже могут принимать участие в формировании ПТП. В этом случае наиболее вероятен электрофоретический механизм перемещения ряда веществ, существующих в ионной форме.
Таким образом, в основе ПТП - длительного увеличения эффективности связи между нейронами - лежат изменения функциональных возможностей пресинаптического волокна.
Посттетаническая потенциация ТПСП тоже может иметь место при высокочастотном раздражении тормозящих афферентных источников (Curtis a. Eccles, 1960). Эффективность тормозящих воздействий при этом будет увеличиваться (Вартанян, неопубликованные данные). Однако данному вопросу уделяется пока мало внимания, и он недостаточно разработан. Следует отметить, что этот вид потенциации будет оказывать на возбудимость нейрона прямо противоположный эффект по сравнению с ПТП возбуждающих синапсов, препятствуя любому повышению возбудимости синаптического происхождения.
От тормозной ПТП следует отличать явления постактивационной депрессии, которая выражается в уменьшении амплитуды ВПСП в постактивационном периоде. Этот вид депрессии многие исследователи связывают с десенситизацией (ослаблением чувствительности к медиатору) субсинаптической мембраны (Thesleff, 1955; Katz a. Thesleff, 1957; Axelsson a. Thesleff, 1958, и др.). Десенситизация мембраны может быть причиной медленного нарастания посттетанической потенциации (Lloyd, 1949) при длительном высокочастотном раздражении, а также объяснять случаи ранней депрессии и относительный характер потенциации. Правда, на нервно-мышечном соединении лягушек и крыс показано, что небольшое число импульсов не уменьшает чувствительности субсинаптической мембраны к медиатору (Otsuka, Endo a. Nonomura, 1962). Но пока нет оснований переносить эти данные на центральные синапсы. Все же более вероятной представляется другая точка зрения, согласно которой ранняя депрессия и относительная потенциация связаны с различным соотношением расхода и мобилизации медиатора (Curtis a. Eccles, 1960).
Следует отметить, что пресинаптическое торможение также является одним из видов пресинаптической депрессии. Оно снижает эффективность возбуждающих синапсов, вызывая деполяризацию пресинаптических окончаний. Обладая противоположным эффектом по сравнению с ПТП, пресинаптическое торможение отличается от нее также и тем, что реализуется через аксо-аксональный синапс (см. выше, гл. 3).
Функциональная роль пресинаптических изменений эффективности синапсов остается еще недостаточно ясной. Но она несомненно велика. Имеется немало оснований считать, что приспособительный характер рефлекторной деятельности в значительной мере обусловливается описанными изменениями эффективности синаптической передачи, которые в настоящее время принято обозначать как пластические свойства синаптического аппарата. Более широким понятием является пластичность нервной деятельности. Она может быть связана не только с пре-, но и с постсинаптическими механизмами, например, с механизмами регуляции возбудимости (см. ниже, гл. 6). Однако наиболее важные элементы, лежащие в основе пластичности, в частности, сохранение следов прошедших возбуждений, по-видимому, тесно связаны с пластичностью синаптических процессов.
Формирование новых связей в ц. н. с. и их закрепление, механизмы хранения следов (памяти) в деятельности нервных центров в значительной мере остаются загадкой. Пока еще не создано нейрофизиологической теории поведения, хотя уже выдвинуто немало гипотез для обоснования наиболее сложных проявлений деятельности мозга.
В последнее время большая роль в формировании "нервного следа" отводится синаптическим образованиям (Eccles, 1964; Eccles a. Meulders, 1964) и биохимическим изменениям в нейронах (Hyden, 1959, и др.).
Ряд исследователей (Konorski, 1948, 1950; Hebb, 1949; Young, 1951; Eccles, 1953, 1957, 1964; Jung, 1953; Mclntyre, 1953, и др.) видят в пресинаптических изменениях эффективности синаптического действия возможную основу памяти и обучения, полагая, что в синапсах могут происходить длительные изменения тонкой структуры под влиянием их "употребления" или "неупотребления". Дж. Экклс с соавторами (Eccles a. Mc Intyre, 1953; Eccles, Krnjevic a. Miledi, 1959) показали, что при перерезке задних корешков (до спинального ганглия) или мышечного нерва за несколько недель до опыта ("неупотребление") моносинаптические рефлексы и ВПСП на поврежденной стороне значительно ослабляются, а потенциационная способность соответствующих пресинаптических волокон значительно возрастает, и ПТП может длиться в течение нескольких часов. Таким образом, при "неупотреблении" существенно изменяются какие-то внутренние свойства синаптических контактов. В частности, может происходить сморщивание пресинаптического волокна, уменьшение пуговок (Eccles a. Mc Intyre, 1953). Связь между степенью функциональной нагрузки и морфологическими изменениями была подтверждена и в ряде других работ (De Robertis a. Franch, 1956; Базанова и др., 1964).
Попытки показать повышение эффективности синапсов при "усиленном употреблении" (Beranek a. Hnik, 1959; Kozak a. Westerman, 1961) пока не дали положительных результатов (R. Eccles, Kozak a. Westerman, 1962; Hnik, Beranek, Vyklycky a. Zelena, 1963).
К сожалению, все эти эксперименты проведены на спинном мозге, где память и обучаемость вряд ли могут быть хорошо выражены. Правда, имеется мнение, что между спинномозговыми нейронами и нейронами высших этажей ц. н. с. нет качественных различий, однако даже сторонники этой точки зрения не отрицают значительных количественных различий в развитии определенных свойств синаптического аппарата нейронов высших и низших отделов ц. н. с. (Eccles, 1964). В частности, потенциация гораздо более выражена у пресинаптических окончаний аксонов пирамидных нейронов по сравнению со спинномозговыми (Land-gren, Phillips, Porter, 1962).
Не исключено, что в высших отделах ц. н. с. повышенная деятельность синаптических полей нейронов оставляет в соответствующих синаптических группах определенный след, так что вероятность активации нейрона с "употреблявшегося" синаптического поля резко возрастает. Возможно, именно этот механизм лежит в основе образования условных рефлексов, проторения возбуждения или торможения по определенным нейрональным цепочкам. С другой стороны, редкая активация синаптического аппарата может в конечном итоге привести к его "самовыключению".
Имеются высказывания и о структурных основах пластичности нервной деятельности. Дж. Экклс (Eccles, 1953), в частности, указывал на возможный рост и набухание пресинаптических окончаний; М. Ф. Купарадзе (1965) отметила способность аксональных окончаний образовывать дополнительные амебовидные отростки.
Наибольшие трудности для "пресинаптической гипотезы" механизмов памяти и обучения состоят в том, что изменения эффективности синапсов в результате "употребления" или "неупотребления" формируются очень длительное время, тогда как условные рефлексы вырабатываются, как правило, после нескольких сочетаний условного и безусловного раздражения. С другой стороны, "пресинаптическая гипотеза" не позволяет объяснить механизмы долговременной памяти, а также принципов организации избирательного облегчения одних связей и угнетения других (Милнер, 1960; Купер, 1961) при формировании условного рефлекса.
Эти трудности пытается преодолеть X. Хиден (Hyden, 1959), выдвигая биохимическую гипотезу механизмов памяти и обучения, сущность которой была изложена выше в главе 1. Соображения Хидена побудили многих исследователей заняться проблемой связи между активностью нейрона и состоянием цитоплазматической РНК. Так, Морелл (Morell, 1963) показал, что в нейронах вторичного эпилептического очага повышается содержание РНК, которая концентрируется преимущественно вдоль внутренней поверхности мембраны. Мэй Чжэнь-тун (1963) обнаружил, что после введения рибонуклеазы выработанные условные рефлексы ослабляются или вовсе исчезают и в дальнейшем не восстанавливаются, хотя могут быть образованы вновь. Проведенный гистохимический анализ выявил уменьшение РНК в нейронах коры головного мозга и полосатого тела. Показано также, что при различных функциональных состояниях в нейронах изменяется и клеточная ДНК (Коган, 1962, 1964).
Химическую гипотезу пластичности и памяти подтверждают и нашумевшие опыты на планариях (McConnell, Jacobson a. Kimble, 1959; Corning a. John, 1961), в которых удалось показать передачу "информации" о наличии временной связи через пищу, состоящую из ранее "обученных" планарий.
В последнее время Сцилард выдвинул новую гипотезу "обучения" и пластичности. Автор полагает, что эффективность и пластичность синаптической передачи определяется взаимодействием набора специфических белков пресинаптической мембраны с соответствующим (комплементарным) набором белков мембраны постсинаптического нейрона. Активные участки пре- и субсинаптической мембраны могут сближаться, обеспечивая физический контакт обеих белковых групп. Реакция рассматривается как аналогичная реакции связывания комплемента (Szilard, 1964).
Химическая гипотеза пластичности, по-видимому, имеет большие перспективы, особенно если она будет учитывать структурно-функциональные особенности нейронной организации мозга. Дж. Экклс отмечает, что пока гипотеза X. Хидена игнорирует функциональную организацию нервной системы и роль пространственного распределения нервных процессов в мозгу (Eccles, 1964). Свойства пластичности и формирование нервных следов (памяти) должны иметь в своей основе несколько механизмов, и становление новых связей в нервной системе должно проходить несколько последовательных этапов.
Вероятно, вначале доминируют чисто функциональные процессы в виде циркуляции нервных импульсов в замкнутых нейрональных цепях (Rashevsky, 1938, и др.). Их реверберационный характер может обеспечивать многократное повторение активации определенных синаптических полей в течение короткого времени, способствуя фиксации определенной пространственно-временной структуры нервных процессов в соответствующих синапсах многих нейронов (Hebb, 1949; Gerard, 1949; Konorski, 1961). Эта фиксация связана уже, по-видимому, с изменением биохимической и морфологической структуры нейрона (рис. 43).
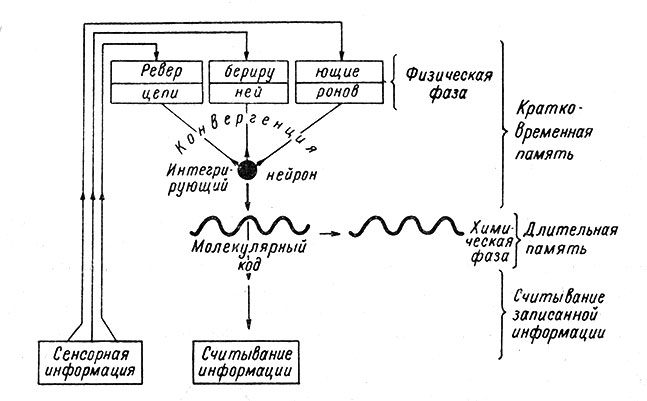
Рис. 43. Гипотетическая схема взаимодействия функционального и химического механизмов памяти. (Маmо, 1962)
Сложное пространственное распределение облегченных и заторможенных связей между нейронами должно быть связано с функциональной архитектурой сложного нервного процесса, протекающего одновременно в нескольких анализаторах. Деятельность анализаторов строится по мнению ряда авторов на принципе формирования нейронной модели нервного стимула, изоморфно отражающей внешний сигнал (Соколов, 1963б, 1964а), и принципа обратной связи, постоянно вносящей коррекцию в распределение активности между отдельными нейронами (Меницкий, 1957, 1964; Анохин, 1962б, 1963). Возникающий на основе этих принципов сложный пространственно-временной рисунок облегченных и заторможенных синаптических полей (в огромном количестве участвующих нейронов) может уже на основании химических и морфологических перестроек фиксироваться на долгое время в мозговых структурах.
Пластические и следовые свойства мозга изучены пока еще недостаточно. Однако уже сейчас очевидно, что пресинаптические изменения эффективности синапсов играют огромную роль в механизмах, лежащих в основе этих свойств.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'