VI. Соотношения между постсинаптическими процессами и импульсной активностью
Как видно из изложенного, возбуждение (ВПСП), торможение (ТПСП и пресинаптическое) и распространяющийся нервный импульс (ПД) имеют неодинаковые ионные механизмы и разыгрываются на разных субстратах. Общим для всех этих процессов является то, что в их основе лежат изменения проницаемости разных участков нейрональной мембраны. Однако на этом их сходство кончается, и начинают доминировать различия. Если это вполне очевидно, когда речь идет о возбуждении (ВПСП) и торможении (ТПСП), поскольку первое характеризуется деполяризацией, а второе гиперполяризацией нейрона, то в случае возбуждения и распространяющегося импульса (ПД) эти различия в известной мере замаскированы тем, что оба процесса характеризуются временной деполяризацией мембраны. Кроме того, в нейрофизиологии традиционно принято говорить о местном (ВПСП) и распространяющемся (ПД) возбуждении.
Действительно, с биофизической точки зрения ПД представляет собой процесс возбуждения по сравнению с состоянием "покоя" нервного волокна (если всякую деполяризацию рассматривать как признак возбуждения). Но следует иметь в виду, что деполяризация, лежащая в основе ПД, отличается от деполяризации при ВПСП не только по физико-химическим механизмам, о чем говорилось выше, но и по ряду кардинальных физиологических свойств.
Известно, что постсинаптическое возбуждение локально, градуально, обладает способностью к суммации, слитно, распространяется с декрементом в силу кабельных свойств мембраны и не оставляет после себя рефрактерное. Нервный импульс является распространяющимся процессом, следующим правилу "все или ничего". Ввиду наличия рефрактерности, связанной с "натриевой инактивацией" и "повышенной калиевой проводимостью" (Hodgkin a. Huxley, 1952a, 1952b), процесс носит дискретный характер, так что отдельные импульсы никогда не сливаются друг с другом и не создают слитного деполяризационного процесса, как в случае постсинаптического возбуждения. При развитии в аксоне местной деполяризации, как в случае пресинаптического торможения, импульсы уменьшаются и могут совершенно исчезнуть при интенсивных деполяризующих токах. Постсинаптическое возбуждение предшествующей деполяризацией не подавляется, а наоборот суммируется с ней.
Различия в функциональных свойствах ВПСП и распространяющегося возбуждения (ПД) были давно хорошо известны нейрофизиологам. Однако наиболее систематически эти сведения были сгруппированы Г. Грундфестом в его работах, посвященных электрической невозбудимости синапсов (Grundfest, 1957, 1959; Грундфест, 1961б, и др.). В них в наиболее отчетливой форме был поставлен вопрос о разделении мембраны нейрона на электрически невозбудимые, но электрогенные участки (субсинаптическая мембрана) и электрически возбудимые участки, или мембраны с проводящим электрогенезом (внесинаптические участки нейрональной мембраны, особенно район начального сегмента аксона и вся аксональная мембрана). Следует отметить, что отличие ионных "каналов", обеспечивающих развитие ВПСП, от "каналов", создающих условия для генерации ПД, в настоящее время рассматривается как общепризнанное (Eccles, 1964).
Но, что самое главное, импульсная активность глубоко отлична от постсинаптического нейронного возбуждения по своему функциональному содержанию. ВПСП в нейроне действительно представляет собой процесс возбуждения в ответ на приходящую по афферентным источникам импульсацию. Это тот самый процесс, который Ч. Шеррингтон в свое время описал на основании косвенных данных и назвал "процессом центрального возбуждения" (Sherrington, 1925). Импульсная же активность лишь передает на последующий нейрон или эффектор в частотно закодированной форме суммарные амплитудные изменения МП нейрона, превышающие критический уровень деполяризации. Импульсная активность отражает общий уровень сверхпороговой деполяризации нейрона, складывающийся в результате всех имеющихся в нем в данный момент возбуждений и торможений (ВПСП и ТПСП). Поэтому импульсная активность в одинаковой степени является "переносчиком" обоих процессов на последующие звенья нейронной цепи. Если нейрон обладает возбуждающими функциями, выделяя возбуждающий медиатор в своих аксональных окончаниях, то конечный эффект при передаче его интегрированного состояния будет носить возбуждающий характер для последующих нейронов. Если же интеграция нервных процессов происходит в тормозящем нейроне, выделяющем в концевых разветвлениях аксона тормозящий медиатор, то его интегрированное состояние будет передаваться на последующее звено как торможение. Таким образом, суммарный деполяризационный процесс в тормозящем нейроне представляет собой основу тормозящих воздействий и функционально является зеркальным отражением ВПСП в возбуждающем нейроне.
С этой точки зрения импульсную активность нельзя отождествлять с постсинаптическим возбуждением. Тогда центральное возбуждение и торможение следует рассматривать как местные процессы, развивающиеся в теле нейронов при активации соответствующих (возбуждающих или тормозящих) синаптических полей. Передача же этих процессов и их взаимодействий осуществляется распространяющимися нервными импульсами, которые характером своего распределения во времени кодируют оба эти процесса в их взаимодействии.
На рис. 37 приведены примеры записей вызванной активности в промежуточном нейроне передних рогов спинного мозга кошки в ответ на совершенно идентичные раздражения заднего корешка L7. Во всех трех приведенных записях ответ нейрона неодинаков. Величина суммарного деполяризационного процесса во всех трех случаях различна, и число вызванных им импульсов неодинаково. По-видимому, к контролируемым воздействиям с заднего корешка прибавляются какие-то неконтролируемые экспериментатором воздействия, которые то увеличивают, то уменьшают деполяризацию. Другими словами, количество возбуждающих и тормозящих влияний во всех трех случаях различно, и эти различные соотношения кодируются различным числом и распределением импульсов, возникающих в нейроне и передающихся по аксону к последующим звеньям.
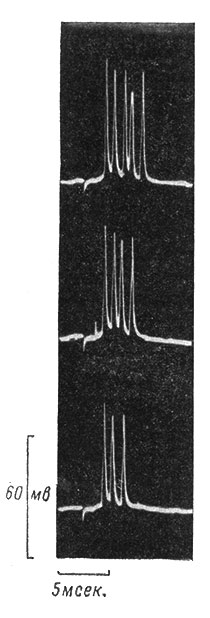
Рис. 37. Вызванные ответы промежуточного нейрона передних рогов спинного мозга кошки на идентичные раздражения заднего корешка L7. Объяснения в тексте
Описанные представления о функциональной роли импульсной активности позволяют понять, почему у И. П. Павлова и его сотрудников были все основания рассматривать возбуждение и торможение как равноценные процессы, с одинаковой скоростью перемещающиеся в высших отделах мозга. Это связано с тем, что импульсная активность, не являясь в информационном смысле ни возбуждением, ни торможением, с одинаковой скоростью передает от нейрона к нейрону как возбужденное, так и тормозное состояние, и служит средством передачи влияний как для возбуждающих, так и для тормозящих нервных клеток.
Можно предполагать, что в ходе эволюции нервная деятельность совершенствовалась в двух основных направлениях: во-первых, по линии развития высокоспециализированных гиперполяризационных реакций, на основе которых формировалось активное координационное постсинаптическое торможение; во-вторых, по линии развития все большего числа диффузных и релейных линий межнейрональных связей с обеспечением быстрой передачи информации по этим линиям от одной группы нейронов к другой в виде помехоустойчивой импульсной активности.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'