V. Ионные механизмы торможения
В настоящее время на нейрональном уровне обнаружено два основных вида торможения: пре- и постсинаптическое. Первое связано с подавлением проведения нервных импульсов в аксональных (пресинаптических) окончаниях афферентных нейронов. Второе обусловливается развитием процесса гиперполяризации в нейронах и получило название тормозного постсинаптического потенциала (ТПСП). TПCП является более оперативным, строго адресованным и универсальным тормозным механизмом и по существу представляет собой то "центральное торможение", которое было постулировано в свое время Шеррингтоном (Sherrington, 1932). Ввиду большой роли, которую играют тормозные постсинантические механизмы в деятельности ц. н. с., а также в связи с тем, что это торможение изучено гораздо полнее и глубже пресинаптического, целесообразно вначале остановиться на ионных процессах, лежащих в его основе.
Развитие ТПСП также связано с изменением проницаемости в так называемых тормозных субсинаптических участках мембраны нейрона. В настоящее время подавляющее большинство нейрофизиологов признает существование двух качественно различных типов синапсов: возбуждающих и тормозящих. Правда, существуют различные точки зрения относительно природы их функциональной специализации. Большая часть исследователей считает, что характер синаптической активации (возбуждающий или тормозящий) связан с химическими свойствами аксональных окончаний пресинаптического (активирующего) нейрона (Eccles, 1964; Шаповалов, 1964). В связи с этим постулируется существование специализированных тормозящих нейронов, аксональные окончания которых выделяют особый медиатор, оказывающий тормозящее воздействие на последующий нейрон. Такой нейрон должен непременно входить в состав тормозящей рефлекторной дуги и сменять возбуждающий медиаторный механизм на тормозящий (Eccles, Fatt a. Landgren, 1956; Eccles, 1961b, 1964). Это предположение подтверждается экспериментальными данными, показывающими, что тормозящее воздействие доходит до изучаемого нейрона на 0.8-1.3 мсек. позже, чем возбуждающее. Данное обстоятельство указывает на дополнительную синаптическую задержку, возникающую вследствие включения вставочного тормозящего нейрона.
Другие исследователи, опираясь на широко известные факты о том, что один и тот же медиатор в различных постсинаптических нейронах может вызывать противоположный эффект (Terzuolo a. Bullock, 1958; Tanc a. Gershenfeld, 1961, 1962, и др.), склоняются к иной точке зрения: по их мнению, различный эффект действия пресинаптических окончаний связан не с тем или другим медиатором, а с особенностями субсинаптических мембран последующего нейрона (Костюк, 1962; Florey, 1962). В последнее время высказывается весьма обоснованное мнение, что, по-видимому, противопоставление этих взглядов носит искусственный характер и что есть достаточно оснований говорить как о "возбуждающей" и "тормозящей" субсинаптической мембране, так и о "возбуждающих" и "тормозящих" окончаниях и медиаторах (Костюк, 1965).
В отличие от изменений проницаемости, происходящих в активированной возбуждающей субсинаптической мембране, в тормозящей мембране при активации происходят более специфические изменения. С помощью тех же экспериментальных приемов, которые использовались для исследования ионных механизмов возбуждения, было выяснено, что в этом случае проницаемость увеличивается только по отношению к некоторым ионам, гидратированные размеры которых не превышают ∼ 2.55 Å (Coombs, Eccles a. Fatt, 1955b; Araki, Ito a. Oscarsson, 1961; Ito, Kostyuk a. Oshima, 1962).
Какие же ионы и в каком направлении должны перемещаться через тормозящую субсинаптическую мембрану, чтобы при этом возникла волна гиперполяризации? Очевидно, общим результатом этих перемещений должно быть относительное увеличение внутри клетки числа анионов. Ток в районе субсинаптической мембраны в этом случае должен быть выходящим (количество отрицательных зарядов внутри нейрона будет при этом возрастать), а в остальных, внесинаптических отделах мембраны - входящим. Проходя через постсинаптическую мембрану, такой ток будет создавать на ней падение напряжения с отрицательным знаком внутри клетки, которое и будет регистрироваться внутриклеточным микроэлектродом в виде ТПСП (рис. 31).
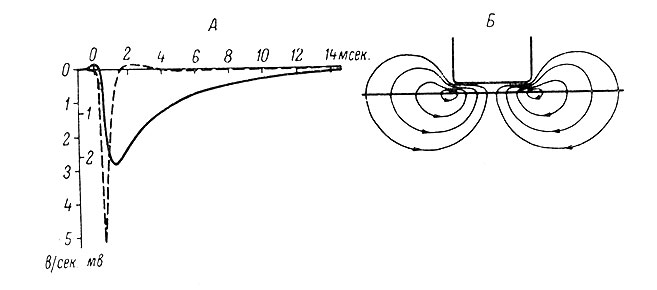
Рис. 31. Развитие во времени ТПСП и генерирующего его тока. (Curtis a. Eccles, 1959). А: сплошная линия - графическое изображение ТПСП, прерывистая линия - графическое изображение расчетной величины постсинаптического тока, генерирующего ТПСП. Б - схематически показаны тормозные постсинаптические токи, генерирующие ТПСП и возникающие в районе тормозных синапсов при активации тормозной субсинаптической мембраны
Уже давно было замечено, что при использовании микроэлектродов, заполненных КСl, в мотонейронах спинного мозга (Coombs, Eccles a. Fatt, 1955b) и в различных других нейронах ц. н. с. (Curtis, Eccles a. Lundberg, 1958; Andersen, Brooks a. Eccles, 1963), включая пирамидные клетки гиппокампа (Kandel, Spenser a. Brinley, 1961; Andersen, Eccles a. L∅yning, 1963), ТПСП из гиперполяризационного быстро превращается в деполяризационный (рис. 32, А, I-III). Из этого факта было сделано заключение, что активированная тормозная субсинаптическая мембрана высоко проницаема для ионов Сl-. Благодаря диффузии из заполненного хлористым калием микроэлектрода значительно повышается внутримембранная концентрация ионов Сl-, в результате чего направление электрохимического градиента для этих ионов меняет свой знак, и выходящий поток ионов Сl- через тормозные зоны мембраны начинает доминировать (рис. 32, Б, В). Поэтому гиперполяризационный ТПСП и превращается в деполяризационный.
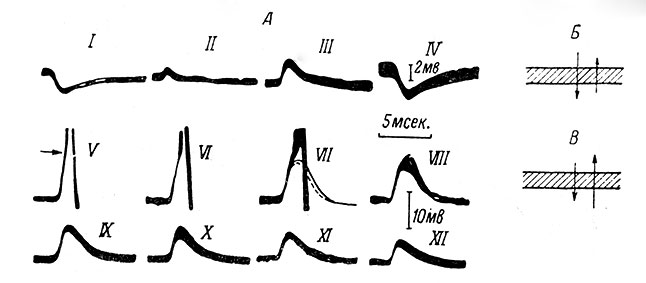
Рис. 32. Превращение гиперполяризационного ТПСП в деполяризационный при поступлении КСl из микроэлектрода внутрь нейрона. А - внутриклеточная регистрация ТПСП, возникающих в мотонейроне m. biceps-semitendinosus в ответ на афферентный залп импульсов в волокнах группы Iа нерва m. quadriceps (Coombs, Eccles a. Fatt, 1955b). Записи произведены с помощью двухканального микроэлектрода, заполненного трехмолярным раствором КСl, и образованы наложением 40 пробегов луча. I-III - влияние диффузии из микроэлектрода ионов Сl- на ТПСП, зарегистрированного при МП покоя (-59 мв); IV - восстановление гиперполяризационного ТПСП при деполяризации мембраны внешним током до низкой величины (-27 мв); V-XII - после пропускания гиперполяризующего тока (силой 3.2 × 10-8 а в течение 60 сек.); все записи зарегистрированы при МП покоя; латентный период потенциалов действия на кривых VI и VII резко различен. Б, В - схематическое изображение потоков ионов Cl- через мембрану до (Б) и после (В) введения этих ионов в составе KCl; на В видно резкое возрастание выходящего потока Cl- (Eccles, 1961a). Внутреннее содержимое клетки - книзу от мембраны. Величина стрелок обозначает интенсивности соответствующих потоков Сl-
Таким образом, участие ионов Сl- в создании ТПСП может считаться несомненным. Однако до последнего времени считалось, что ЕCl равен приблизительно -70 мв (Экклс, 1957, и др.). В то же время изучение величины ТПСП на фоне изменения уровня МП нейрона с помощью пропускания постоянного тока через один из каналов двухканального микроэлектрода показало, что ЕТПСП равен приблизительно -80 мв (Coombs, Eccles a. Fatt, 1955b). Эти данные были подтверждены с помощью методики фиксации МП, когда изучению подвергаются токи, обусловливающие ТПСП (Araki a. Terzuolo, 1962). Рис. 33 иллюстрирует сказанное. Из рисунка также видно, что величина ТПСП меняется в зависимости от уровня МП, увеличиваясь при низких его значениях и уменьшаясь при его приближении к -80 мв; при дальнейшем увеличении МП по абсолютной величине знак ТПСП меняется - он становится деполяризационным.
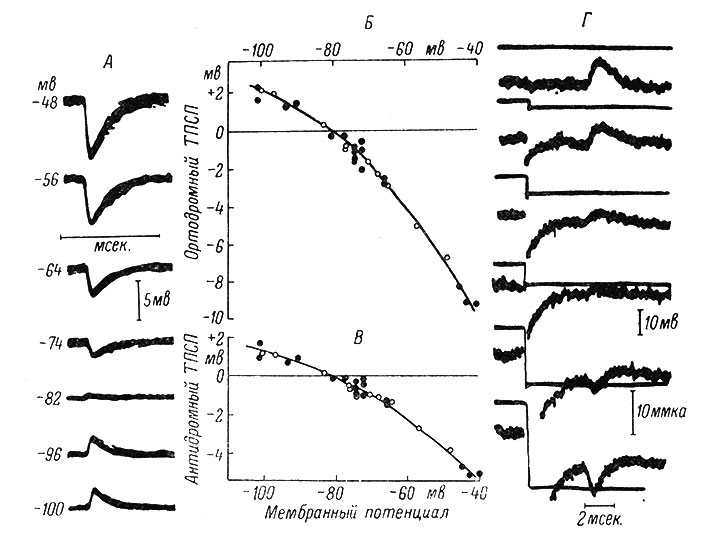
Рис. 33. Зависимость между уровнем мембранного потенциала и величиной ТПСП. Потенциал равновесия ТПСП. А - внутриклеточные записи ТПСП в мотонейроне ядра нерва m. biceps-semitendinosus с помощью двухканального микроэлектрода. Записи, полученные наложением около 40 пробегов луча, показывают ТПСП, вызванные афферентными залпами в n. quadriceps. С помощью постоянного тока, пропускаемого через второй канал микроэлектрода, МП сдвигается до уровней, обозначенных у каждой записи. Исходный МП покоя -74 мв. Б - график измерений записей, частично представленных на А. По оси абсцисс - мембранный потенциал; по оси ординат - соответствующие величины ТПСП. В - график измерений записей ТПСП, вызванного в том же мотонейроне антидромным раздражением; обозначения те же, что и на A (Coombs, Eccles a. Fatt, 1955b). Г - верхние и нижние кривые представляют собой соответственно записи МП и тока, необходимого для фиксации мембранного потенциала на постоянном уровне. Верхняя пара кривых является контрольной. В последующих записях, как это видно по верхним кривым, перед вызовом тормозных постсинаптических токов применялись гиперполяризующие прямоугольные токи, которые длились в течение всей записи (Araki a. Terzuolo, 1962)
Из сказанного вытекает два вывода. Во-первых, очевидно, что при развитии тормозного субсинаптического тока ионы, формирующие этот ток, движутся по своим электрохимическим градиентам. Во-вторых, движение ионов хлора по электрохимическому градиенту не может обеспечить гиперполяризационный ТПСП при величине МП большей, чем -70 мв. В создании ТПСП должен принимать участие еще какой-то ион, потенциал равновесия которого выше потенциала равновесия ТПСП (EТПСП). Таким ионом в нервной клетке является К+.
В опытах по изучению ионных потоков, вызывающих следовую гиперполяризацию после полного ПД мотонейрона, было показано, что она создается за счет повышения калиевой проводимости постсинаптической мембраны и выхода ионов К+ из клетки. Потенциал равновесия следовой гиперполяризации, а следовательно, и ЕK равен приблизительно -95 мв (Coombs, Eccles a. Fatt, 1955a; Kuno, 1959; Araki, Ito, Kostyuk, Oscarsson a. Oshima, 1962). Поэтому можно предполагать, что ионы К+ принимают существенное участие в формировании ТПСП, выходя из нейрона во время активации тормозных субсинаптических мембран и вызывая тем самым гиперполяризацию (Coombs, Eccles a. Fatt, 1955b).
Точно проверить гипотезу об участии ионов К+ в создании ТПСП в нейронах млекопитающих оказалось невозможным. До сих пор экспериментально не удается существенно изменить внутриклеточную концентрацию К+, а следовательно, определить степень его участия в тормозных субсинаптических токах. Пока делается допущение, что К+ принимает в создании ТПСП приблизительно такое же участие, как и Сl-.
Участие К+ в создании гиперполяризационного ТПСП в последнее время подтверждается новыми данными о существовании активного переноса ионов хлора снаружи внутрь (так называемый накачивающий хлорный "насос"). Действие такого "насоса" будет поддерживать внутриклеточную концентрацию на более высоком уровне, чем это необходимо для диффузионного равновесия. ЕСl в связи с этим будет смещен в сторону некоторой деполяризации по отношению к нормальному МП. Следовательно, ионы Сl-, перемещаясь при активации тормозной мембраны по своему градиенту, не смогут вызывать гиперполяризационного процесса. Напротив, их перемещение будет сопровождаться некоторой деполяризацией. Учитывая все это, есть основания считать, что гиперполяризация при ТПСП создается выходящими из нейрона ионами К+ (Eccles, Eccles a. Ito, 1964).
Несмотря на то что ионы хлора при достаточно высоком уровне МП (порядка -70 мв) выходят во время развития ТПСП из клетки и тем самым вызывают ее деполяризацию, потоки хлора все же препятствуют развитию импульсной активности в нейроне. Дело в том, что критический уровень деполяризации значительно ниже ECl. В то же время, как только уровень МП снижается до значений, меньших ECl направление потока ионов Сl- меняется (рис. 32), и хлор начинает поступать внутрь нейрона, гиперполяризуя его или по крайней мере препятствуя дальнейшей деполяризации. Таким образом, повышенная проницаемость к ионам Сl- в тормозных субсинаптических мембранах стабилизирует МП на уровне значительно более высоком, чем критический, и препятствует развитию ПД в нейроне (Eccles, Eccles a. Ito, 1964).
В некоторых тормозных синапсах избирательность проницаемости еще более суживается. В мышечных волокнах ракообразных тормозный эффект возникает только за счет входа в клетки ионов Cl- (Boistel a. Fatt, 1958). В сердечных волокнах ТПСП связан с выходом из клеток ионов К+ (Trautwein, Kuffler a. Edwards, 1956; Trautwein a. Dudel, 1958). Эта суженная избирательность может быть объяснена наличием на стенах "пор" активированной тормозной мембраны положительных или отрицательных зарядов (Boistel a. Fatt, 1958). В первом случае "поры" будут проницаемы только для ионов Сl-, во втором - для ионов К+.
В связи с раскрытием основных закономерностей перемещения ионов при развитии постсинаптического торможения появилась возможность для построения гипотезы о механизме действия ионов брома. Как известно, бром в оптимальных дозах оказывает усиливающее влияние на центральное торможение. Среди многочисленных работ, посвященных изучению действия этого вещества, наиболее демонстративны исследования школы И. П. Павлова, в которых на огромном фактическом материале было показано усиливающее действие препаратов брома на процессы условного торможения. Однако по поводу механизма действия этих веществ высказывались самые различные точки зрения, в том числе предположения о том, что бром оказывает свое влияние через ретикулярную формацию ствола мозга (Крылов, 1959, 1960). И лишь сравнительно недавно появились исследования, показавшие непосредственное влияние брома на деятельность нервных клеток (Бирюков, 1963; Яковлев, 1963; Либерман и Цофина, 1964). В дальнейшем с помощью методики внутриклеточного отведения было показано, что внутривенное введение бромистого натрия (0.05-0.1 г/кг) увеличивает постсинаптическое торможение в мотонейронах спинного мозга кошек (рис. 34), не оказывая никакого влияния на ВПСП (Вартанян, Яковлев и Александрова, 1964). Последующий анализ показал, что нет оснований приписывать действие брома пресинаптическим механизмам, поскольку он не увеличивает активности клеток Реншоу - тормозящих промежуточных нейронов (Вартанян, Яковлев, Александрова, 1966), осуществляющих так называемое возвратное торможение мотонейронов спинного мозга. Поэтому можно предполагать, что механизмы действия брома на ТПСП имеют постсинаптическое происхождение.
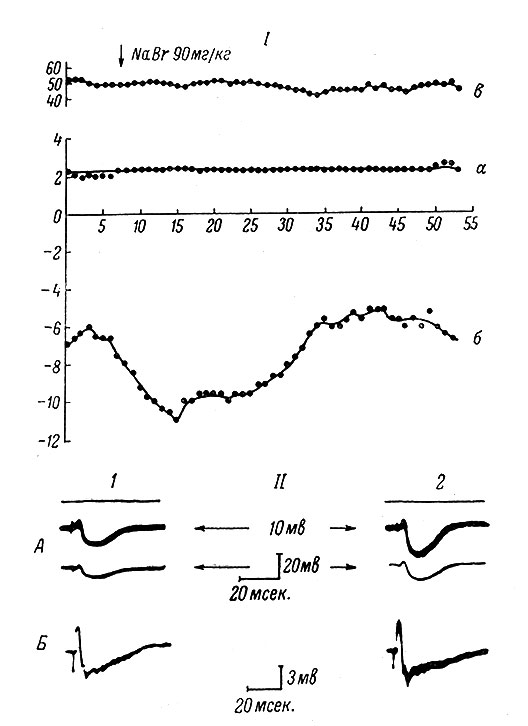
Рис. 34. Влияние бромистого натрия на постсинаптическое торможение (собственные данные). I. По оси ординат - величины ВПСП (а) и ТПСП (б) и МП (в) в период записи комбинированных (ВПСП + ТПСП) постсинаптических потенциалов; по оси абсцисс - время (в мин.) после введения микроэлектрода в мотонейрон и начала записи комбинированных постсинаптических потенциалов, возникающих на раздражение заднего корешка L7 повторными стимулами (длительность 0.1 мсек., частота - 0.5/сек.; сила - 5 порогов моносинаптического ВПСП). Каждая точка представляет собой соответственно величину ВПСП или ТПСП в комбинированном ответе, записанном путем наложения 20 одиночных пробегов луча. Стрелка - момент введения 90 мг/кг NaBr. II. А - внутриклеточные записи комбинированного постсинаптического потенциала до (2) и через 10 мин. после (2) внутривенного введения NaBr в дозе 90 мг/кг. Раздражается задний корешок L7. Параметры раздражения даны в подписи к I. Нижний луч по постоянному току одновременно записывает уровень мембранного потенциала. Калибровка для нижнего луча 20 мв, для верхнего луча (по переменному току) - 10 мв. Вверху отмечена нулевая линия. Записи образованы путем наложения 20 одиночных пробегов луча. Б - аналогичные записи в другом мотонейроне до (1) и через 10 мин. после (2) внутривенного введения NaBr в дозе 170 мг/кг. Раздражается задний корешок L6. Условия и параметры раздражения те же, что и на А. Запись по переменному току
Как близкий к Сl галоген, ион Вr- должен вступать во все ионные трансмембранные процессы. Если процесс активного переноса Сl- извне во внутрь клетки (Eccles, Eccles а. Itо, 1964) специфичен только по отношению к этому аниону, то ЕВr- может быть сдвинут относительно ECl- в сторону гиперполяризации, что может обеспечивать большую величину тормозной гиперполяризации при введении ионов Br- в организм. Определенное значение могут иметь при этом и меньшие размеры этого иона по сравнению с ионами Cl-, обеспечивающие большую его проходимость через тормозящую субсинаптическую мембрану, что было показано в недавних исследованиях с внутриклеточной электрофоретической инъекцией различных анионов (Araki, Ito a. Oscarsson, 1961; Но, Kostyuk a. Oshima, 1962).
Таковы в общих чертах основные закономерности перемещения ионов в период активации тормозящих субсинаптических мембран. В основе этих перемещений, как и в случае возбуждения, лежат изменения проницаемости определенных участков нейрональной мембраны. Различия заключаются только в степени увеличения этой проницаемости, т. е. в степени увеличения "пор" в субсинаптической мембране, что приводит к участию различных ионов в трансмембранных потоках и в свою очередь к различным изменениям уровня поляризации клетки. Более специфический характер изменений проницаемости в тормозной субсинаптической мембране по сравнению с изменениями в возбуждающей позволяет думать о том, что развитие тормозных механизмов является относительно более поздним эволюционным приобретением. Действительно, к деполяризационным процессам, лежащим в основе процесса возбуждения, способны самые разнообразные живые клетки. Деполяризация не является специализированной реакцией и не представляет собой привилегии нервных клеток. Активный гиперполяризационный процесс является высокоспециализированной реакцией нервных и некоторых мышечных клеток, которая, надо полагать, развилась вторично в процессе эволюции, в результате чего в нервной системе возникли условия для сложнейших координации (Орбели, 1950; Бирюков, 1960).
Впервые убедительные доказательства в пользу существования пресинаптического торможения были получены Франком и Фьюортесом в 1957 г. (Frank a. Fuortes, 1957). В дальнейшем этот вид торможения был подробно изучен в лаборатории Дж. Экклса большой группой исследователей, и в настоящее время основные сведения о нем изложены в ряде обобщающих работ (Eccles, 1963, 1964, 1965; Костюк, 1965).
Имеются два основных отличия пресинаптического торможения от постсинаптического. Во-первых, все события при развитии пресинаптического торможения разыгрываются в так называемых аксоаксональных синапсах, в которых субсинаптической структурой является не тело клетки, а пресинаптические разветвления афферентного волокна (Gray, 1962, 1963; Szentagothai, 1963, и др.). Во-вторых, основным электрофизиологическим феноменом этого торможения является деполяризация аксональных окончаний.
Экспериментально показано, что предварительное ("кондиционирующее") раздражение волокон группы Iа, идущих от аннулоспиральных окончаний, вызывает пресинаптическое торможение первичных афферентных волокон той же группы. Раздражение волокон Ib оказывает тормозящее влияние на те же волокна мышц-антагонистов (флексоры влияют на экстензоры), а также на волокна флексорной группы Iа и кожные первичные афферентные волокна (Eccles, Schmidt a. Willis, 1936b). Общая схема пресинаптических влияний разных групп волокон на первичные афферентные окончания в спинном мозге представлена на рис. 35.
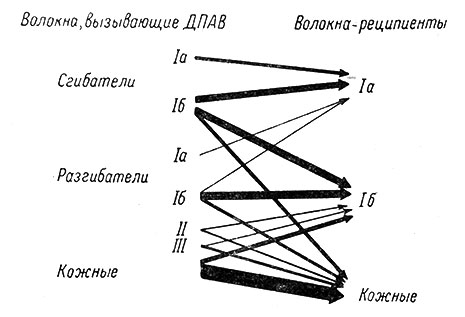
Рис. 35. Схема деполяризующих влияний различных афферентных волокон на волокна групп Iа, Ib и кожные волокна α. (Eccles, Schmidt a. Willis, 1963b). Слева указаны виды афферентных волокон, которые вызывают деполяризацию первичных афферентных волокон (ДПАВ). Ширина стрелок дает представление о величине ДПАВ, возникающей в волокнах-реципиентах
Никаких изменений МП постсинаптического нейрона под влиянием пресинаптического торможения не происходит. Оно вызывает лишь уменьшение величины ВПСП в мотонейронах на протяжении 200-300 мсек. (Frank a. Fuortes, 1957; Frank, 1959; Eccles, Eccles a. Magni, 1961). В настоящее время экспериментально доказано, что это уменьшение ВПСП, а следовательно, и уменьшение рефлекторного разряда, связано с деполяризацией пресинаптических волокон (Eccles, Eccles a. Magni, 1961; Eccles Magni a. Willis, 1962; Eccles, Schmidt a. Willis, 1962, и др.).
Пресинаптическое торможение выявляется приблизительно через 5 мсек. после нанесения кондиционирующего раздражения, достигает максимума через 20 мсек. и длится, как правило, дольше 200 мсек. (Eccles, Eccles a. Magni, 1961, и др.). Учитывая длительность центральной синаптической задержки (около 0.5 мсек.), можно полагать, что центральная часть пути пресинаптического торможения состоит не менее чем из двух последовательно соединенных промежуточных нейронов. В промежуточном ядре были обнаружены нейроны, обладающие соответствующими свойствами (Eccles, Kostyuk a. Schmidt, 1962a). Значительная длительность деполяризации первичных афферентных волокон (ДПАВ), лежащей в основе пресинаптического торможения, может быть связана с длительным действием вызывающего ее медиатора или с большой постоянной времени мембраны (Eccles, 1964). Однако не исключено, что существенное значение здесь играют промежуточные нейроны, вызывающие эту деполяризацию, которые могут быть соединены в структуру, осуществляющую длительную циркуляцию возбуждения по замкнутому кругу и соответственно длительное его действие на пресинаптические окончания (Костюк, 1965).
Ионные механизмы пресинаптического торможения, точнее говоря ДПАВ, лежащей в его основе, практически не изучены из-за невозможности ввести микроэлектрод в достаточно тонкие пресинаптические волокна. Несомненно, что ДПАВ возникает в результате изменения проницаемости мембраны пресинаптических волокон при выделении медиатора в аксоаксональных синапсах. Природа этого вещества неизвестна, однако оно не идентично медиатору, вызывающему постсинаптическое торможение. Стрихнин, полностью блокирующий последнее (Bradley, Easton a. Eccles, 1953; Fatt, 1954; Экклс, 1957; Curtis, 1959, и др.), не оказывает заметного влияния на пресинаптическое торможение (Eccles, 1962; Eccles, Schmidt a. Willis, 1963d; Schmidt, 1963). В то же время оно достаточно полно блокируется пикротоксином.
Можно полагать, что неизвестный медиатор повышает проницаемость мембраны ко всем ионам, как в случае ВПСП, и что поскольку при этом возникает деполяризация, то ионы Na+ должны занимать существенное место в суммарном ионном потоке. Однако определить потенциал равновесия ДПАВ пока еще не удалось. Предполагается, что он сдвинут от уровня МП в сторону деполяризации по крайней мере на 30 мв (Eccles, Schmidt a. Willis, 1963a). Искусственная гиперполяризация, так же как и следовая гиперполяризация афферентных волокон после тетанизации задних корешков, всегда увеличивают ДПАВ (Eccles a. Krnjevic, 1959; Eccles, Magni a. Willis, 1962; Eccles, Schmidt a. Willis, 1963a). Деполяризация афферентных волокон уменьшает ее, однако даже большие деполяризационные токи (до 0.6 мА) не приводят к реверсии ДПАВ. А только ее реверсия была бы доказательством того, что в основе ДПАВ лежит ионный механизм с определенным потенциалом равновесия.
Таким образом, пока имеются лишь качественные данные о соответствии между величиной деполяризации афферентных пресинаптических волокон и степенью подавления их синаптической эффективности. Хорошей иллюстрацией этой закономерности являются данные о соотношении пассивной деполяризации этих волокон и величиной вызываемых ими ВПСП, полученные Экклсом, Костюком и Шмидтом (рис. 36). Как видно из рисунка, деполяризация первичных афферентных волокон сопровождается уменьшением ВПСП, а их гиперполяризация вызывает его увеличение.
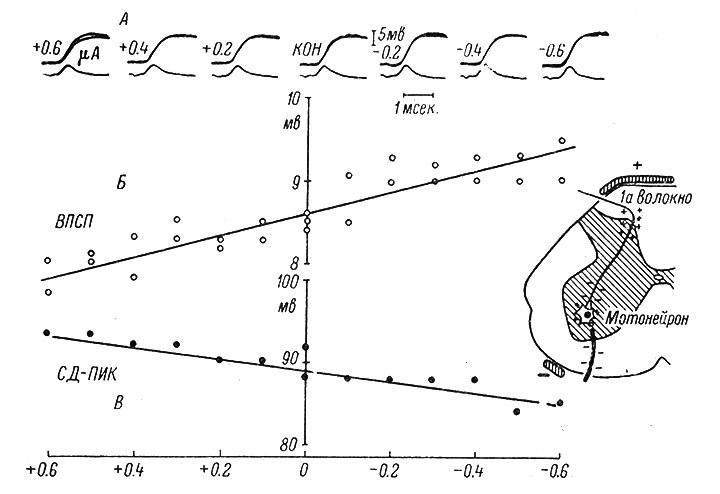
Рис. 36. Изменения моносинаптических ВПСП при пропускании поляризующего тока через спинной мозг кошки. (Eccles, Kostyuk a. Schmidt, 1962c). Внутриклеточные записи от мотонейрона, иннервирующего m. biceps anterior; мембранный потенциал -70 мв. А - примеры записей ВПСП при нарастании поляризующего тока обоих значений; величины тока (в мка) указаны рядом с каждой записью; в середине - контрольная (исходная) величина ВПСП. Каждая запись состоит из ряда наложенных друг на друга одиночных пробегов луча. Верхние записи - внутриклеточные, электрически продифференцированные на нижних записях. Калибровка дана только для недифференцированных записей. Б - амплитуды ВПСП, частично представленных на А, отложены на графике. По оси абсцисс - величина и направление поляризующего тока; по оси ординат - амплитуда ВПСП в мв. В - изменение амплитуды антидромно вызванного потенциала действия (СД-пик) под влиянием тех же поляризующих токов. На А, Б, В (+) и (-) указывают на полярность дорсального электрода. На вставке - схема поляризации и возникающего распределения зарядов в различных участках афферентных волокон и мотонейронов
Как же влияет ДПАВ на прохождение афферентной импульсации в пресинаптических волокнах? Деполяризация пресинаптических волокон приводит к уменьшению величины проходящих через них ПД. В точных опытах на гигантском синапсе кальмара было показано количественное соотношение между величиной ПД пресинаптического волокна и амплитудой ВПСП в постсинаптическом волокне (Hagiwara a. Tasaki, 1958; Takeuchi a. Takeuchi, 1962). Оказалось, что логарифм величины ВПСП находится в прямой линейной зависимости от амплитуды пресинаптического ПД (Eccles, 1964). Это означает, что даже небольшое уменьшение амплитуды ПД приводит к резкому снижению величины ВПСП в постсинаптическом нейроне. По-видимому, аналогичные изменения ВПСП при снижении пресинаптического ПД имеют место и в спинном мозгу (Eccles, 1964).
Несколько слов о функциональном значении пресинаптического торможения. Этот вид торможения у млекопитающих распространен в основном в синаптическом аппарате первичных волокон спинномозговых афферентных систем, так что поступление интенсивной стимуляции по одной из них может снижать эффективность импульсации в других системах или даже полностью блокировать их деятельность. Пресинаптическое торможение оказывает влияние не только на собственный рефлекторный аппарат спинного мозга, но и влияет на первичные синаптические реле ряда восходящих к головному мозгу трактов (Eccles, Kostyuk a. Schmidt, 1962b; Eccles, Schmidt a. Willis, 1963c). Известно также нисходящее пресинаптическое торможение первичных афферентных окончаний волокон группы Iа и кожных афферентов (Andersen, Eccles a. Sears, 1962, 1964; Carpenter, Lundberg a. Norrsell, 1962; Carpenter, Endberg a. Lundberg, 1962; Andersen, Eccles, Schmidt a. Yokota, 1963a, 1963b, и др.).
В спинном мозгу пресинаптическое торможение часто выступает в роли своеобразной отрицательной обратной связи, ограничивающей афферентный поток при сильных раздражениях и, таким образом, в известной мере выполняет охранительную функцию по отношению к спинномозговым центрам. Интенсивное афферентное раздражение может затормаживать все остальные афферентные потоки и, по-видимому, создавать условия для возникновения доминантного очага возбуждения, обеспечивающего реализацию наиболее "срочных" и "важных" рефлексов.
Нисходящее пресинаптическое торможение, очевидно, является первым "ярусом" активного ограничения поступающей извне информации и может рассматриваться как пример "перцептивного внимания" (Eccles, 1965), обеспечивающего влияние высших центров на поступающие в ц. н. с. потоки раздражений.
В высших отделах головного мозга, в частности в коре больших полушарий, доминирует постсинаптическое торможение (Eccles, 1964), которое обеспечивает тонкие аналитико-синтетические функции высших отделов мозга.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'