I. Особенности биохимии мозга
1. Высокая скорость обмена и преобладание аэробных процессов. Как показали проведенные Кети и его сотрудниками (Kety, 1952, 1962) измерения на людях in vivo, средняя величина потребления кислорода головным мозгом составляет 3.3 мл О2 на 100 г мозговой ткани в 1 мин. Если принять средний вес мозга за 1400 г, то общее потребление кислорода всем мозгом составит 46 мл/мин. Интересно, что эта величина весьма постоянна: при сне и бодрствовании, при вдыхании кислорода и углекислого газа, при введении гормонов, при шизофрении и других психических заболеваниях значительно меняется лишь кровоток через мозг, тогда как потребление кислорода остается неизменным. Лишь при диабетической и в особенности при инсулиновой коме выявлено снижение потребления кислорода мозговой тканью до 1.7-1.9 мл О2/100 г/1 мин. (Kety, 1962).
Если принять объем дыхательного воздуха за 500 мл, частоту дыхания за 16 в 1 мин., а содержание О2 во вдыхаемом и выдыхаемом воздухе соответственно за 21 и 17%, то общее потребление О2 телом человека составит 320 мл в 1 мин., т. е. только в 7 раз больше, чем потребление одним мозгом (табл. 1).
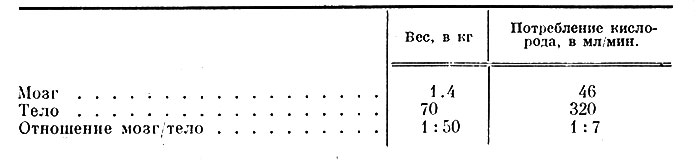
Таблица 1. Потребление кислорода мозгом и телом человека (Kety, 1962)
2. Преимущественное потребление глюкозы крови. Хотя за последние годы выявлено наличие в мозгу гликогена и активное участие его в функционально-биохимических превращениях в мозговой ткани (Прохорова, 1960), общее содержание в мозгу гликогена (70 мг%), а также глюкозы (45 мг%) крайне мало. Поэтому основным источником энергии для мозговой ткани является глюкоза крови: мозг человека, например, потребляет в 1 мин. в среднем 75 мг глюкозы (Kety, 1962). Простой расчет показывает, что это составляет 108 г глюкозы в сутки; энергия от сгорания такого количества глюкозы равна 450 ккал, т. е. 1/6-1/8 от суточных энергетических трат организма человека.
3. Богатство липидами. Для мозговой ткани характерно высокое содержание различных видов липидов, в особенности - фосфолипидов. За последние годы удалось измерить абсолютную скорость обновления этих соединений: она оказалась весьма высокой (табл. 2); по-видимому, липиды обновляются в целом со скоростью, близкой к скорости обновления белковых молекул мозга.

Таблица 2. Время полного обновления фосфолипидов мозга
Показано, что липиды мозга вовсе не являются инертным, чисто пластическим материалом: их обмен активно меняется при различных функциональных состояниях нервной системы (Крепс, 1956; Heald, 1960; Rossiter, 1962).
4. Альтернативные пути обмена. Для мозговой ткани весьма характерно то, что одно и то же соединение может расщепляться до конечных продуктов двумя разными, обычно взаимоисключающими путями, с образованием разных промежуточных продуктов (Tower, 1963). Так, α-кетоглутаровая кислота может превращаться в янтарную непосредственно через сукцинил-КоА, а может вначале переаминироваться с γ-аминомасляной кислотой в полуянтарный альдегид, и уже последний окисляется до янтарной кислоты. Существование альтернативных путей показано и для пировиноградной кислоты. По-видимому, возможность переключения обмена то на один, то на другой путь обеспечивает в какой-то степени ту быстроту биохимических изменений, которая должна соответствовать достаточной скорости смен физиологических состояний нейрона.
5. Компартментализация обмена веществ. Под этим несколько громоздким термином понимают наличие в нервной ткани по крайней мере двух пространственно разделенных фракций одного и того же вещества, причем меньшая фракция характеризуется более высокой скоростью обмена, основная же фракция обменивается значительно медленнее. Например, активная фракция глутаминовой кислоты в мозгу, составляющая около 20% всей глутаминовой кислоты мозга, имеет период полужизни менее 1 часа, а основная фракция - период порядка 6-8 час. (Waelsch, 1959; Waelsch et al., 1964). Такая компартментализация (от слова compartment - отделение, отсек, купе и т. п.) доказана для глутаминовой кислоты, глутамина, аммиака, γ-аминомасляной кислоты, α-кетоглутаровой кислоты, глицина и других компонентов азотистого обмена (Waelsch a. Lajtha, 1961; Lajtha, 1964). Существование двух фракций вещества - активной и резервной - наблюдается и в других органах; однако в печени, например, почти вся глутаминовая кислота находится в активной форме, в отличие от мозга, где эта форма составляет лишь небольшую часть общей массы глутаминовой кислоты (Berl et al., 1962). По-видимому, явление компартментализации имеет большое значение именно для осуществления быстрой функциональной перестройки обмена веществ в нервной ткани.
6. Гемато-энцефалический барьер. Гисто-гематические барьеры существуют в той или иной форме во всех органах, но гемато-энцефалический барьер несомненно является наиболее сложным. Сущность этого барьера, несмотря на большое число посвященных ему работ, до сих пор не ясна. Совершенно очевидно, что гемато-энцефалический барьер осуществляет активный перенос веществ из мозга в кровь и из крови в мозг, причем скорость первого процесса в целом выше, чем второго. В то же время для отдельных аминокислот (глутаминовая, γ-аминомасляная кислота, тирозин) показано, напротив, избирательное поглощение из крови (Tower, 1963; Udenfriend, 1963; Lajtha, 1964). Несомненно, что общая скорость проникновения вещества в мозг зависит как от исходных химических свойств молекулы данного вещества, так и от его участия в динамике биохимических процессов мозга.
7. Роль медиаторов. Эти низкомолекулярные биологически активные соединения являются тонкими регуляторами обменных процессов в нервной ткани. При этом они, вероятно, участвуют как в регуляции постоянно текущих процессов обмена веществ, так и в особенности в экстренной биохимической перестройке нейрона при развитии возбуждения и торможения. Обе эти функции медиаторов - трофическая и пусковая - подверглись за последние годы детальному анализу. Однако несмотря на большое число исследований по биохимической активности в мозгу ацетилхолина (Nachmansohn, 1959; Мс Intosh, 1963; Демин, 1963), катехоламинов (Утевский и Осинская, 1963; Fugazza, 1963), серотонина (Costa, 1960), γ-аминомасляной кислоты (Владимиров и Сытинский, 1961), гистамина (White, 1961), механизм биохимического действия медиаторов все еще не ясен.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'