Глава XIV. Историческое развитие тканей
Мы показали ранее, что становление многоклеточное было сопряжено с дифференцировкой тела на два органа или две ткани - кинобласт и фагоцитобласт, обеспечивавшие самые элементарные функции живого - отграничение от внешней среды, обмен веществ, чувствительность или раздражимость и движение. Клетки в пределах одной ткани имели одинаковую дифференцировку, в разных - различную. Причем уже на самой заре эволюции многоклеточных животных филогенетическая дифференциация фагоцитобласта происходила неодинаково.
У кишечнополостных, от которых впоследствии произошли все Deuterostomia (вторичноротые животные, в том числе позвоночные, включая человека), он поляризовался и тело их оказалось состоящим из двух эпителиобразных тканевых слоев - наружной эктодермы и внутренней энтодермы. У червей, давших начало Protostomia (первичноротым), он более длительное время оставался паренхиматозным и лишь позднее в нем произошла дифференциация на поляризованный слой клеток, выстилающий просвет образовавшегося кишечника - центральный фагоцитобласт и рыхлую паренхиму - периферический фагоцитобласт. Таким образом, в результате вторичной поляризации клеток возникла вторая пограничная ткань - эпителий кишечного типа - энтодермальный.
В периферическом фагоцитобласте шла дальнейшая филогенетическая дифференциация клеток (рис. 65А). В связи с увеличением размеров тела животного возникла потребность в опорных структурах и среди клеток периферического фагоцитобласта дифференцировались десмобласты - клетки, с которыми было связано образование волокнистых и мембранных структур. Остальные клетки остались амебоцитами. Так появилась соединительная ткань. Следовательно, на этом этапе филогенетического развития тело многоклеточного животного было представлено тремя функциональными системами клеток, образующими три тканевых типа - эктодермальный эпителий, энтодермальный эпителий и ткань внутренней среды.
Аналогичная тканевая дифференцировка происходила и во второй основной ветви многоклеточных животных - у вторичноротых.
На протяжении дальнейшей эволюции, в результате уже третичной поляризации клеток внутренней среды, из периферического фагоцитобласта возникла пограничная ткань, образованная плоскими клетками, выстилающими вторичные полости тела (брюшную, грудную, околосердечную) - мезотелий. Шла дифференциация и в пределах ткани внутренней среды: возникли кровь, хрящевая и костная ткани.
Появление соматической мышечной ткани и нервных элементов можно представить себе как результат дифференциации кинобласта, в котором совмещались функции отграничения, локомоторная (двигательная) и восприятия раздражений. По-видимому, среди клеток эктодермы появились клетки, специализировавшиеся в выполнении обеих последних функций (рис. 65Б), - они прежде и чувствительнее других клеток пласта воспринимали раздражения из внешней среды и осуществляли, благодаря развившимся в них сократительным волоконцам, более совершенную локомоцию, чем с помощью ресничек.
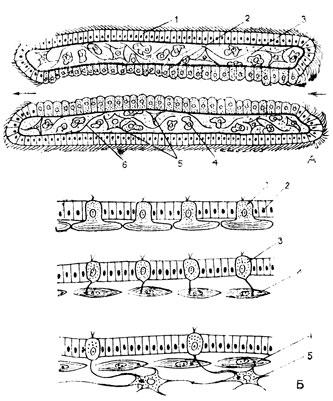
Рис. 65. Предполагаемая филогенетическая дифференцировка периферического фагоцитобласта (Л) и кинобласта (Б). А: 1 - эктодерма; 2 - энтодерма; 3 - формирующаяся ткань внутренней среды; 4 - амебоцит; 5 - десмобласты; 6 - мембранные и волокнистые структуры. Б: 1 - эпителиально-мышечная клетка; 2 - эктодермальная клетка; 3 - чувствительная клетка; 4 - мышечная клетка; 5 - нервная клетка
Позднее, в результате дальнейшего разделения и интенсификации функций, произошло филогенетическое "расщепление" такой чувствительно-двигательной клетки на две еще более специализированные - чувствительную и двигательную. Последние в совокупности образовали соматическую мышечную ткань. Наконец, часть чувствительных клеток погрузилась во внутреннюю среду организма и, дифференцировавшись в многоотростчатые невроны, образовала первичную, диффузную, сетевидную нервную систему. Остальные первичные чувствительные клетки сохранили синаптические связи с мышечными клетками и установили новые с невронами.
В итоге помимо пограничных возникли функциональные системы клеток, выполняющих специализированно двигательную, сократительную и чувствительную функции. То есть в результате филогенетически закрепленной функциональной дифференцировки клеток возникли мышечная и нервная ткани.
Постановка и разработка проблемы закономерностей и путей эволюции тканей - заслуга советской гистологии. Особенно интенсивно работа над этой проблемой протекала в 30 - 40-х годах, когда возникли два различных направления, две школы - А. А. Заварзина и Н. Г. Хлопина.
Вопросы эволюционной гистологии разрабатывались отдельными зарубежными и отечественными учеными и раньше, но первым, кто глубоко занялся проблемой эволюции тканей и своими замечательными работами привлек к ней внимание гистологов, был академик А. А. Заварзин. Правда, ранние его исследования были посвящены выявлению чисто формальных морфологических закономерностей. Они были оторваны от идеи филогенеза и поэтому носили статический (неподвижный) характер. Однако в последующих работах А. А. Заварзина эта первоначальная теоретическая ошибка была преодолена и проблему, которой были посвящены его дальнейшие исследования, он назвал уже "эволюционной динамикой тканей". Здесь им решались вопросы о движущих силах тканевой эволюции, о специфических закономерностях исторического развития тканей.
К концу жизни и творчества А. А. Заварзина его учение об эволюции тканей приобрело вполне материалистический характер. Основными установками этого учения явились: 1) единство формы и функции в развитии при ведущей роли функции; 2) взаимодействие организма со средой; 3) целостность, интегрированность организма и коррелятивные взаимосвязи его тканевых систем. Теория А. А. Заварзина в ее последних формулировках построена на понимании глубокой связи структуры с функциональными изменениями частей организма, вызванными его адаптивной, приспособительной эволюцией, протекающей в единстве с окружающей средой. Тканевая эволюция как эволюция элементарных систем организма, лишенных качества органической индивидуальности, обусловливается согласно данной теории эволюцией целых организмов - происходит естественный отбор организмов по мутационным изменениям клеток тканей, наилучшим образом обеспечивающим их функционирование в данных условиях существования.
А. А. Заварзин обратил особенное внимание на черты сходства в строении тканей совершенно неродственных групп животных и считал характерной чертой эволюции тканей принцип их параллельного и конвергентного1 развития в филогенезе. Напомним для примера сходство в строении поперечнополосатой соматической мышечной ткани у позвоночных и членистоногих, в строении хряща у позвоночных и головоногих моллюсков, наконец, сходство в тонком строении ткани внутреннего скелета позвоночных - кости и наружного скелета насекомых - хитина. Сходство в строении аналогичных тканей у неродственных животных обусловлено, по А. А. Заварзину, одинаковостью выполняемых этими тканями функциональных задач в организме. Параллелизм морфологических изменений в тканях возникает в силу того, что при огромном разнообразии условий существования организмов и не меньшем количестве адаптивных изменений в строении животных, функциональные запросы самых различных организмов к своим тканям довольно однообразны. Эти функциональные запросы А. А. Заварзин сводил к четырем основным функциям: пограничности (включая обмен с внешней средой), внутреннего обмена, двигательной и нервной. Таким образом, даже в разнообразных условиях существования самым различным организмам требуется от тканей одно и то же, и эволюция этих тканей происходит одинаково - в направлении повышения их рабочей производительности.
1 (Конвергенция - сходство признаков, обусловленное общностью функции.)
Основоположник другого направления по проблеме тканевой эволюции академик Н. Г. Хлопин исследовал, наоборот, черты различия в строении как гомологичных1, так и аналогичных тканей. Ведущим в определении природы тканей он считал генетический момент. Историческое развитие тканей согласно теории Н. Г. Хлопина подчинено общим морфологическим закономерностям дивергентной2 эволюции.
1 (Гомологичные - родственные по своему происхождению в филогенезе.)
2 (Дивергенция - расхождение признаков.)
Таким образом, существуют теория параллелизма гистологических структур А. А. Заварзина и теория дивергентной эволюции и филогенетической системы тканей Н. Г. Хлопина.
Что же, однако, лежит в основе тканевой эволюции - принцип дивергенции или же принцип конвергенции и параллелизма гистологических структур?
А. А. Заварзин в монографии "Очерки эволюционной гистологии крови и соединительной ткани" значительное место уделяет вопросу дивергенции тканевых элементов и тканевых систем. В то же время Н. Г. Хлопин всегда признавал наличие широко представленных параллелизмов в строении тканей. Однако каждый из этих ученых считал ведущим в историческом развитии тканей именно выдвигаемый им принцип. По нашему же с В. П. Михайловым (А. А. Браун и В. П. Михайлов, 1958, 1961, 1966) глубокому убеждению, решение вопроса о закономерностях и путях тканевой эволюции лежит в творческом синтезе обеих этих теорий. Согласно нашим взглядам, каждая из упомянутых теорий отражает лишь одну сторону процесса тканевой эволюции. В эволюционном развитии тканей выявляется единство противоположностей - конвергенции, а также параллелизмов и дивергенций. Эволюция тканей, как нам представляется, осуществляется на основе параллелизма их дивергентных изменений.
В процессе исторического развития организмов дивергируют все категории их признаков, в том числе и тканевые структуры. По ходу филогенеза возрастает многообразие тканей. Однако все это многообразие укладывается, в основном, в 4 тканевые группы, отвечающие четырем элементарным свойствам живых систем - пограничности, внутреннего обмена, раздражимости и сократимости. Ткани сак строительные материалы проще и функционально беднее чем образуемые ими органы - конструкции, и поэтому они отвечают более однообразной перестройкой на сходные запросы (перестройкой, основанной на отборе через целостные организмы мутационных изменений в клеточных элементах их тканей). Вот почему в строении тканей параллелизмы выражены заметнее и чаще, чем в строении органов и надерганных частей тела.
Пути эволюции тканей, намеченные на основе развиваемых нами представлений, показаны на схеме (рис. 66). Схема эта ориентировочная, носит характер грубого наброска, но и в таком, так сказать, первом приближении достаточно наглядно отражает нашу основную мысль. Схема состоит из системы линий, которые при сопоставлении их друг с другом, с одной стороны, как бы дивергируют, расходятся, а с другой стороны, обнаруживают при этом расхождении параллельное расположение, то есть они одновременно оказываются и расходящимися, и параллельными, что и должно графически иллюстрировать наше представление о параллелизмах в дивергентном развитии тканей.
Пути филогенетического развития тканей начинаются на нашей схеме с гипотетической Паренхимеллы или фагоцителлы И. И. Мечникова. Все они заканчиваются в одной из четырех колонок, отвечающих четырем основным группам: во-первых, тканям первичным - пограничной и внутренней среды, уже наметившихся у фагоцителлы (соответственно кинобласта - К и фагоцитобласта - Ф), и, во-вторых, тканям вторичным - мышечной и нервной.
Дифференцировка тела фагоцителлы на два слоя - наружный (кинобласт) и внутренний (фагоцитобласт), иначе говоря на два органа и в то же время на две ткани, была обусловлена ее взаимоотношением с внешней средой: наружный слой выполнял прежде всего ограничительную, защитную функцию; внутренний слой - функцию внутреннего обмена. Обмен с внешней средой осуществлялся через кинобласт - пласт живых клеток, принимающих активное участие в этом обмене. Поначалу этим же пограничным клеточным пластом одновременно выполнялась и функция восприятия раздражений из внешней среды и функция движения. В процессе исторического развития кинобласт начало разным видам кожного, эпидермального эпителия (Э-1), ткани нервной системы (Н-1) и соматической мышечной ткани (М-1).
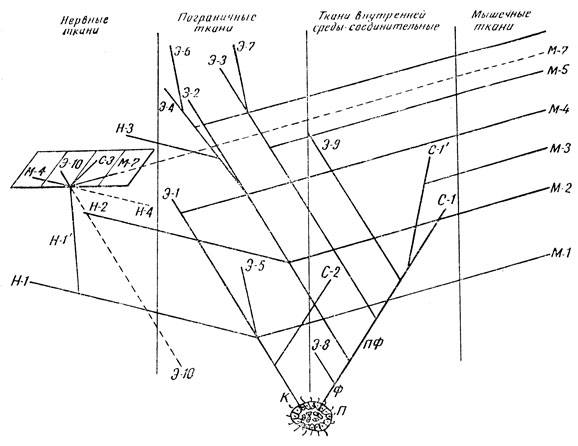
Рис. 66. Пути тканевой эволюции (по А. А. Брауну, 1958). П - паренхимелла, или фагоцителла Мечникова. К - кинобласт; Ф - фагоцитобласт; ПФ - периферический фагоцитобласт; Э-1- кожный, эпидермальный эпителий; Э-2- кишечный, энтеродермальный эпителий (поляризованный центральный фагоцитобласт); Э-3- целомический, целодермальный эпителий артикулят и моллюсков; Э-4 - целомический, целодермальный эпителий вторичноротых; Э-5 - выделительный, нефродермальный эпителий червей; Э-6 - выделительный нефродермальный эпителий позвоночных; Э-7 - выделительный, нефродермальный эпителий моллюсков; Э-8- эпителиобразный покров губок; Э-9- эндотелий позвоночных; Э-10 - десцеметов эпителий роговицы глаза, пигментный эпителий сетчатки, эпителий мозговых оболочек, эпендимная выстилка спинномозгового канала; С-1 и С-1'- соединительная ткань; С-2 - эктодермальные производные, принимающие участие в образовании мезоглии; С-3 - микроглия; Н-1 - нервная ткань эктодермального происхождения; Н-1' - невроглия позвоночных; Н-2 - нервная ткань кишечной нервной системы кишечнополостных (энтодермальное нервное сплетение) и субэпителиального нервного сплетения глотки и кишки иглокожих (энтоневральный отдел нервной системы); Н-3 - нервная ткань гипоневрального отдела нервной системы иглокожих; Н-4 - фоторецепторные элементы сетчатки глаза - клетки-палочки и клетки-колбочки; М-1 - соматическая мышечная ткань, включая эпителиально-мышечные клетки эктодермы кишечнополостных и наружного покрова турбеллярий; М-2 - эпителиально-мышечные клетки энтодермы кишечнополостных; М-3 - внутренностная мышечная ткань позвоночных; М-4 - миоэпителиальные элементы позвоночных; М-5 - целомическая мышечная ткань насекомых (средней кишки); М-6 - сердечная целомическая мышечная ткань позвоночных, М-7 - мионевральные элементы радужины глаза позвоночных
Из ныне живущих многоклеточных три наиболее примитивные группы: губки, кишечнополостные и низшие безкишечные ресничные черви обладают телом, расчлененным на кинобласт и фагоцитобласт. У губок, как приспособление к сидячему образу жизни, усиливался ток пищевых частичек из внешней среды внутрь тела путем образования жгутиковых камер за счет пограничного пласта клеток; в результате кинобласт у взрослых животных оказывается расположенным внутри фагоцитобласта. Покровная же функция перешла к фагоцитобласту, на поверхности которого дифференцировался слой эпителиобразно залегающих пинакоцитов (Э-8). Наоборот, у Antozoa, высших из кишечнополостных Cnidaria, мезоглия (ткань между двумя пластами пограничных клеток - эктодермой и энтодермой) обладает значительной толщиной и состоит из студенистого основного вещества, иногда волокнистого строения. В образовании этой ткани внутренней среды принимает участие кинобласт путем миграции в нее групп эктодермальных клеток (С-2).
В процессе эволюционной дифференцировки первичный фагоцитобласт дал, в свою очередь, начало двум тканям -1) центральному фагоцитобласту, то есть эпителию кишечного типа (Э-2), возникшему в результате вторичной поляризиции клеток, расположенных во внутренней среде организма, и 2) аполярной мезенхиме, послужившей источником развития всего многообразия соединительных тканей (С-1 и С-11). Причем интересно отметить, что поляризация центрального фагоцитобласта, приводящая к образованию кишечника, происходила независимо, параллельно у кишечнополостных и у ресничных червей.
В процессе дальнейшей эволюции возникла вторичная полость тела, или целом, выстланная собственным эпителием. Целом возник независимо, с одной стороны, у кольчатых червей и моллюсков путем частичной поляризации уже периферического фагоцитобласта, а с другой стороны, у вторичноротых (к числу которых относятся хордовые) энтероцельным путем, то есть из кишечника (центрального фагоцитобласта). Параллельное образование целодермального эпителия показано на схеме линиями Э-3 и Э-4. Точно так же, независимо друг от друга, произошла дифференцировка выделительного, почечного, нефродермального эпителия в разных группах животных. Он возник параллельно у кольчатых червей из эпидермального эпителия (Э-5), у позвоночных (Э-6) и моллюсков (Э-7) из эпителия целодермального.
Дифференцирующийся в системе тканей внутренней среды позвоночных эндотелий (Э-9) приобретает конвергентно, в силу сходства функций, большое морфологическое сходство с эпителием целомической выстилки - мезотелием.
Очень демонстративно независимое, параллельное образование мышечной ткани в процессе дивергентного развития, то есть эволюционной дифференцировки других тканей. Так из кинобласта возникла соматическая мускулатура (М-1), включая эпителиально-мышечные клетки эктодермы кишечнополостных. Аналогичным образом развились эпителиально-мышечные клетки энтодермы (М-2) этих же животных. В системе тканей внутренней среды позвоночных дифференцировалась гладкая внутренностная мышечная ткань (М-3). У позвоночных животных за счет эпидермального эпителия возник миоэпителий (М-4) кожных желез. Из целомической выстилки насекомых развились мышцы их средней кишки (М-5), целомический эпителий позвоночных дал начало сердечной мышечной ткани (М-6).
Наконец, в процессе филогенеза происходило параллельное и независимое развитие нервной ткани из различных эпителиальных пластов. Из ныне живущих форм кишечнополостные и иглокожие обладают, наряду с эктоневральной (Н-1) и энтоневральной (Н-2) нервной системой, возникшей из кишечного эпителия, но построенной из таких же гистологических элементов, что и нервная система, развившаяся из кинобласта (Н-1). У иглокожих имеется еще третий отдел нервной системы (Н-3), развившийся из целомического эпителия.
Особенный интерес представляет процесс филогенетической дифференцировки невроглии (Н-11) позвоночных. Обслуживая невроны, невроглия выполняет в пределах нервной системы самые различные функции - пограничную, трофическую, опорную. В случае надобности на нее ложится и сократительная функция. В особых условиях ее элементы несут и функцию восприятия раздражений. Претерпевая дифференцировку, приспосабливаясь к выполнению этих разных задач, неироглия дивергирует в различных направлениях. Однако эволюционное развитие невроглии не выходит за пределы все тех же четырех морфофункциональных групп, Таким путем в составе невроглии возникли параллельно, независимо от других участков тела тканевые структуры пограничного характера (Э-10) - десцеметов эпителий роговицы, пигментный эпителий сетчатки, эпителий мозговых оболочек и эпендимная выстилка спинномозгового канала. В процессе филогенетической дифференциации появились клетки звездчатой формы с волокнистыми структурами, выполняющие опорную и упаковочную функцию в нервной системе (астроцитарная глия, С-3). Развившиеся в радужине мышечные элементы (М-7), гладкие у млекопитающих и поперечнополосатые у рептилий и птиц, являются мионевральными элементами, конвергентно приобретшими чрезвычайное сходство, соответственно с внутренностной и соматической мышечной тканью позвоночных. Клетки-палочки и клетки-колбочки сетчатки глаза (Н-4), которые академик А. А. Заварзин относил к первым невронам оптического пути, на основании тщательных экспериментальных исследований Я. А. Винникова должны рассматриваться в качестве особой свето- и цветочувствительной модификации эпендимы.
В настоящем кратком очерке и схеме мы не касаемся деталей вопроса о путях тканевой эволюции и вовсе не претендуем на полноту данной картины. Мы поставили здесь лишь задачу показать на ряде важнейших примеров, что в основе филогенетического развития тканей лежит закономерность параллелизма их приспособительных морфофункциональных дивергентных изменений.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'