Глава XI. Мышечные ткани
Мышечные ткани, в отличие от опорно-трофических, представляют собой сборную группу. Они разнятся друг от друга не только по морфофункциональным признакам, но, главное, по своей природе и происхождению как в фило-, так и в онтогенезе. Важнейшими тканями этой группы являются у позвоночных: 1) гладкая внутренностная мышечная ткань, 2) поперечнополосатая скелетная, или соматическая мышечная ткань, 3) сердечная мышечная ткань. Сюда же должны быть отнесены миоэпителиальная и мионевральная ткани позвоночных, а также поперечнополосатая, гладкая соматическая мышечная и эпителиально-мышечная ткани беспозвоночных.
Гладкая мышечная ткань позвоночных близка по своей природе тканям внутренней среды и, как все последние, развивается из эмбриональной мезенхимы. Это клеточная ткань. Ее структурные и функциональные единицы - гладкие мышечные волокна представляют собой клетки, чаще всего веретенообразной формы (рис. 53А). Несравненно реже, например, в эндокарде сердца или в мочевом пузыре лягушки встречаются многоотростчатые мышечные клетки. В последнем случае мышечные волокна очень удлинены, а их отростки иногда напоминают нити. Природа гладкомышечных клеток, близкая основным клеточным элементам внутренней среды - фибробластам, проявляется в способности их к коллагенообразованию. Эти клетки объединяются в ткань прежде всего при помощи коллагеновых футляров, одевающих их тело, а также коллагеновых перемычек между этими футлярами (рис. 53Б). И то, и другое является непосредственным продуктом деятельности самих гладких мышечных клеток. Фибробласты имеются лишь в прослойках рыхлой соединительной ткани между группами гладкомышечных волокон. По этим прослойкам в мышечную ткань входят сосуды и нервы. В ней залегают и все другие клеточные и волокнистые элементы, присущие соединительной ткани. Особенно много эластических волокон, как антагонистов сократительных элементов. Образуя ткань, гладкие мышечные волокна располагаются плотно и своими заостренными концами входят в промежутки между такими же концами других мышечных клеток.
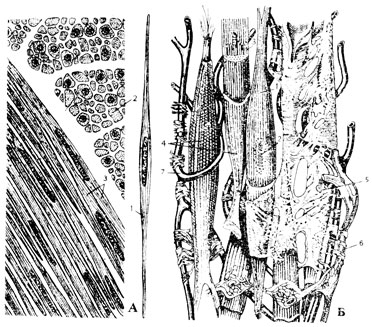
Рис. 53. Гладкая мышечная ткань. А - изолированная гладкая мышечная клетка (1) и гладкомышечная ткань в продольном (3) и поперечном (2) разрезах. Б - схема строения гладкой мышечной ткани по Крелингу и Грау: 1 - гладкая мышечная клетка; 2 - ее ядро; 3 - миофибриллы; 4 - коллагеновый футляр; 5 - эндомизий; 6 - нерв; 7 - кровеносный капилляр
Ядра у гладкомышечных клеток (гладких миоцитов) имеют вид более или менее коротких палочек и лежат в утолщенной части клетки несколько эксцентрично. Рядом с ядром располагается клеточный центр. Помимо органоидов общего характера, в гладких мышечных клетках имеются органоиды специального значения - сократительные мышечные волоконца, миофибриллы, образующие сократительный аппарат клетки. Миофибриллы под световым микроскопом кажутся однородными, гомогенными, почему их и назвали гладкими, а содержащие такие миофибриллы клетки получили наименование гладких мышечных. Миофибриллы имеют толщину 1 - 2 мк и расположены вдоль клетки, по ее периферии. С помощью электронного микроскопа было установлено, что сократительные волоконца имеют сложное строение и состоят из тончайших протофибриллей - гладких миофиламентов диаметром 100 Å.
Гладкая мышечная ткань входит, главным образом, в состав внутренних органов - в толщу стенок органов пищеварительного тракта, воздухоносных путей, сосудов и пр. Из нее построены также залегающие в коже мышцы, поднимающие волосы и перья. Поднятая дыбом при виде собаки шерсть кошки - результат сокращения таких мышц. Тем же объясняется и часто наблюдаемое на холоду нахохливание воробьев. Сокращение гладких мышечных волокон непроизвольное, не зависит от воли человека и регулируется так называемым вегетативным, или внутренностным отделом нервной системы. Так же непроизвольно происходит перистальтическое, волнообразно бегущее вдоль кишечника сокращение мышечной оболочки его стенки, продвигающее вперед пищевые массы. Другой физиологической особенностью деятельности гладкой мышечной ткани позвоночных является медленность сокращения ее волокон и их малая утомляемость.
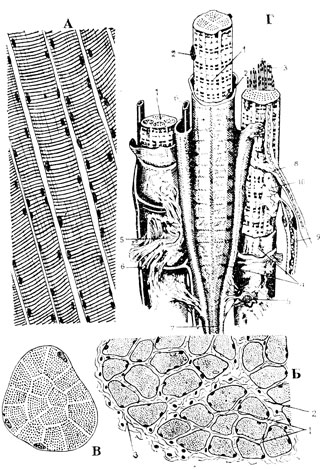
Рис. 54. Поперечнополосатая скелетная мышечная ткань. А - ее продольный разрез; Б - поперечный разрез: 1 - эндомизий; 2 - перимизий; 3 - эпимизий. В - поперечный разрез мышечного волокна (видны столбики, группы миофибриллей). Г - схема строения поперечнополосатой мышечной ткани по Крелингу и Грау: 1 - мышечные волокна (мион); 2 - ядро миона; 3 - миофибриллы; 4 - сарколемма; 5 - эндомизий; 6 - кровеносные капилляры; 7 - сухожильная нить; 8 - вегетативное нервное волокно; 9 - соматическое (двигательное) нервное волокно; 10 - концевая моторная бляшка
Скелетная поперечнополосатая мышечная ткань позвоночных развивается из миотомов. Она противоположна гладкой по своим физиологическим свойствам - волокна ее сокращаются быстро и по воле человека, они более или менее быстро утомляемы. Эта ткань - активный компонент аппарата движения, главная составная часть мускулов тела. Она представлена так называемыми мионами (рис. 54), надклеточными образованиями симпластического характера (симпласт - тело, образованное общей протоплазматической массой со многими ядрами внутри).
Волокна соматической мышечной ткани могут достигать 10 см в длину и 70 мк в толщину. Их удается видеть и невооруженным глазом. Тонкие нити, на которые легко распадается хорошо разваренное мясо, как раз и представляют собой отдельные мионы. В длинном поперечнополосатом мышечном волокне, точнее в его саркоплазме, залегает большое количество ядер; иногда число их измеряется многими десятками. Вблизи ядер встречаются канальцы гранулярного ретикулума и элементы пластинчатого комплекса.
По всей длине волокна проходят миофибриллы, обнаруживающие поперечную исчерченность. Толщина каждой миофибриллы составляет всего 0,5 - 2 мк. Она состоит из чередующихся темных и светлых участков - дисков. Темные диски обладают двойным лучепреломлением и называются анизотропными, или А-диска-ми (рис. 55). Светлые диски носят название изотропных, или И-дисков. Так как во всех миофибриллах совпадают уровни залегания соответственно А-дисков и И-дисков, волокно в целом приобретает правильную поперечную полосатость.
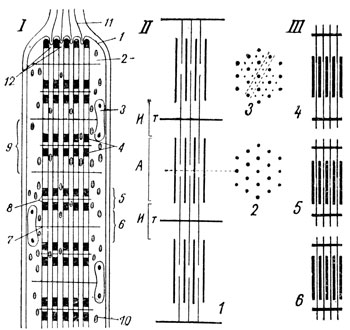
Рис. 55. Схема строения поперечнополосатого мышечного волокна. I - микроскопическое строение волокна: 1 - сарколемма; 2 - саркоплазма; 3 - ядро; 4 - миофибриллы; 5 - темный диск (А); 6 - светлый диск (И); 7 - телофрагма; 8 - мезофрагма; 9 - саркомер; 10 - митохрндрии; 11 - сухожильные пучки; 12 - впячивания плазмолеммы. II - субмикроскопическое строение миофибриллы. Значения А, И, Т - те же. 1 - видны два типа миофиламентов - толстые миозиновые протофибриллы и тонкие, актиновые. При проведении среза так, как указано на схеме 3, между толстыми протофибриллами были бы видны по две тонких. III - положение миофиламентов в разных состояниях мышечного волокна: 4 - в покое; 5 - при неполном и 6 - при полном сокращении
Электронномикроскопическое изучение показало, что миофибриллы поперечнополосатых волокон состоят из протофибриллей - миофиламентов двух видов. Одни из них более толстые, 100 Å и более в диаметре, залегают в области А-дисков. Другие более тонкие, 40 - 60 Å в толщину, располагаются на большем своем протяжении в области И-дисков, но концами заходят с обеих сторон на территорию А-дисков. Между двумя толстыми миофиламентами, построенными из белка миозина, вставлены, в зависимости от того, как прошел срез, один или два тонких, состоящих из белка актина. Суммарно выделяемый из мышц сократимый белок является актомиозином.
Мышечное волокно с поверхности одето сарколеммой (по-гречески "сарко" означает мясо), образуемой совместно саркоплазмой и окружающей мион рыхлой соединительной тканью. Внутренняя часть сарколеммы представлена плазмолеммой, снаружи к ней тесно прилежат волокнистые структуры. На месте связи с сухожилием (на конце мышечного волокна) плазмолемма образует внутрь саркоплазмы глубокие впячивания, за которыми следуют анатомически в глубь миона, но не соприкасаясь непосредственно с его саркоплазмой, пучки коллагеновых волокон сухожилия.
Мышечные волокна пронизываются на равных расстояниях телофрагмами-перегородками, пересекающими миофибриллы через середину светлых дисков (рис. 56). Этими телофрагмами каждый И-диск как бы делится на две равные части. Их обозначают условно буквами T или Z. Участок от одной телофрагмы до другой получил наименование саркомера, иначе - мышечного сегмента. Телофрагма в переводе на русский язык означает конечная перегородка. В мионах имеются еще и другие перегородки - мезофрагмы (М), проходящие через середину темных дисков.
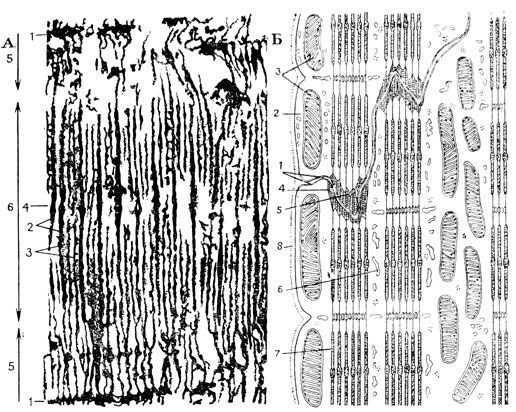
Рис. 56. Электронномикроскопическое строение поперечнополосатой мышечной ткани. А - электронограмма миофибриллы скелетной мышечной ткани (по Хаксли): видны толстые (2) и тонкие (3) протофибриллы светлого, изотропного диска - 5; телофрагмы - 1; мезофрагма - 4. Участок фибриллы от 1 до 1 - саркомер. Б - пограничная область двух соседних клеток сердечной мышцы (из Баргмана): 1 - плазмолеммы двух соседних клеток; 2 - саркоплазма; 3 - митохондрии; 4 - вставочная пластинка с закрепленными в ней миофиламентами (5); 6 - канальцы эндоплазматической сети; 7 - миофиламенты; 8 - сарколемма
В отличие от телофрагм, они в саркоплазму за пределы миофибрилл не входят (рис. 55 и 56). Как выяснилось с помощью электронномикроскопического исследования, при сокращении мышечного волокна происходит вдвигание тонких актиновых протофибриллей между толстыми миозиновыми навстречу друг другу. Этим и объясняется резкое уменьшение в сократившемся мионе И-дисков. В то же время на месте телофрагм наблюдаются полосы сокращения. Создается впечатление, что при максимальном сокращении мышечного волокна часть материала из актиновых нитей перетекает в телофрагмы.
На акт мышечного сокращения расходуется много энергии. Ее поставщиком в мионе являются довольно многочисленные здесь митохондрии, получившие название саркосом. Они отличаются сильным развитием крист. Саркосомами особенно богаты высокоактивные мионы.
Наконец, благодаря электронному микроскопу была открыта в мышечных волокнах особая саркоплазматическая сеть - разновидность гладкого ретикулума в виде канальцев, расположенных между миофибриллами и образующих вздутия около телофрагм. Были обнаружены также тонкие трубковидные впячивания сарколеммы, залегающие в саркоплазме около телофрагм и параллельно им, то есть поперек волокна.
Среди скелетных мышечных волокон различают красные и белые по соотношению между количеством саркоплазмы и миофибрилл. В красных меньше сократительных волоконец, в белых они почти нацело выполняют мионы и оттесняют ядра на периферию к сарколемме. Белые мышечные волокна - это мионы более сильного сокращения, но зато и более утомимые. В мускулах человека мозаично представлены обе разновидности мионов. В чистом виде белые мышечные волокна имеются в грудных мускулах у кур. В хвосте рыбы - основном органе, обеспечивающем ее поступательное движение, мионы белые. В мускулах, с сокращением которых связана слабая, но непрерывная работа грудных плавников, поддерживающих тело рыбы в определенном положении, мионы красные. В таком красном мышечном волокне сократительные волоконца иногда образуют пучок незначительного размера, располагающийся эксцентрично в обильной саркоплазме.
Мионы объединяются в мускулы с помощью рыхлой соединительной ткани, по которой проходят сосуды и нервы (рис. 54). Наиболее тонкие прослойки, непосредственно связанные с сарколеммой, образуют эндомизий; группы мышечных волокон, одетые более толстыми соединительноткаными прослойками - перимизий, а вокруг всего мускула залегает эпимизий, содержащий иногда более или менее значительные скопления жировых клеток.
Мионы скелетной мускулатуры, будучи в сформированном состоянии симпластами, развиваются из клеток. В недавнее время этот вопрос был переисследован с применением электронной микроскопии и предложена новая схема данного процесса. Согласно новым представлениям процесс развития миосимпластов начинается у эмбриона с деления премиобластов (рис. 57), которые дают начало первому поколению миобластов. Каждый из таких миобластов делится митотически и дает 2 клетки. Одна из дочерних клеток - миобласт второго поколения в свою очередь делится, а вторая клетка остается в интерфазе. В результате еще трех последовательных делений из одного миобласта второго поколения возникают 8 миобластов пятого поколения. Собираясь в цепочку, а затем сливаясь друг с другом, эти 8 миобластов строят мышечную трубку (еще незрелое мышечное волокно со сравнительно небольшим количеством миофибрилл и поэтому с центральным расположением ядер).
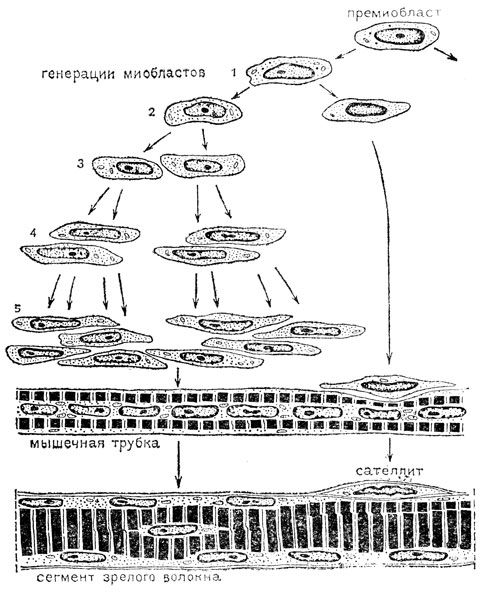
Рис. 57. Схема предполагаемого миогенеза от премиобласта до сегмента мышечного волокна с сателлитом (по Чург)
В это время вторая дочерняя клетка миобласта первой генерации, задержавшаяся в интерфазе, прикладывается к мышечной трубке. Далее количество миофибрилл в волокне нарастает, и они оттесняют ядра к периферии. Поверх зрелого мышечного волокна образуется микроскопически видимая оболочка - внутренний слой сарколеммы. Малодифференцированная дочерняя клетка миобласта первой генерации оказывается под данной оболочкой. Таким клеткам дали название сателлиты, или миосателлиты.
Таким образом, поперечнополосатое волокно скелетной мускулатуры оказывается состоящим из сегментов, непосредственно переходящих друг в друга. Каждому сегменту отвечают 8 ядер дифференцировавшихся миобластов пятой генерации и одна камбиальная клетка - сателлит. Под электронным микроскопом в месте прилегания сателлита к мышечному волокну видно, что каждое из этих образований одето собственно плазмолеммой.
С сателлитами связывают процесс самообновления мышечной ткани и ее репарации после повреждений.
У членистоногих мионы поперечнополосатой мышечной ткани построены еще более сложно, чем у позвоночных, и период мышечной исчерченности волоконец содержит больше участков, хотя структурный принцип миофибрилл остается тем же и их основными элементами являются актиновые и миозиновые протофибриллы, объединяющиеся при сокращении в актиномиозиновый комплекс. Большая сложность строения мионов взаимообусловлена высоким совершенством их функции. Виртуоз скрипач, двигая пальцами, может проделать около 10 сокращений мышц в секунду, а колоратурная певица достигает в пределе 25 сокращений мышц, колеблющих голосовые связки. В то же время обычная муха способна сокращать мышцы крыла сотни раз в секунду.
Однако у других беспозвоночных, например, у червей или моллюсков соматические мышечные волокна резко отличаются от мионов позвоночных и человека прежде всего гладким характером их миофибрилл. Эти мышцы могут быть очень сильными, например, запирательная мышца у двустворчатых мягкотелых. Но сокращаются они медленно и соответственно их миофибриллы гладкие.
Сердечная мышечная ткань позвоночных развивается из участка спланхнотома. Как и соматическая, она является поперечнополосатой (рис. 56Б), поскольку представляет собой ткань быстрого сокращения. Однако она малоутомима и данное обстоятельство находится в связи с красным характером ее волокон. Как известно, ее сокращения не зависят от нашей воли. Микроскопической особенностью сердечной мышечной ткани является сетевидное объединение ее волокон - параллельно расположенные волокна соединяются друг с другом перемычками, анастомозами. Вторая особенность этой ткани - наличие по ходу волокон особых перегородок, вставочных пластинок, значение которых было неясно. А. А. Заварзин рассматривал их как внутриволоконные сухожилия, способствующие синхронному сокращению всей мышечной сети сердца.
Электронный микроскоп показал, что волокна сердечной мышечной ткани представляют собой цепочки одно-двуядерных клеток-миоцитов, а вставочные пластинки - толщенные границы между ними. Сердечные миоциты - крупные клетки, достигающие в длину 120 мк. Миофибриллы не переходят из клетки в клетку, а закрепляются в области вставочных дисков-пластинок. Места крепления получили название десмосом, т. е. соединяющих телец. Клетки сердечной мышцы богаты саркоплазмой и поэтому ядро не оттесняется на периферию, а лежит в центре. Миофибриллы же занимают периферическое положение. Толщина миофибрилл 1 - 3 мк. Их структура такая же, как и в мионах - они состоят из сотен толстых миозиновых тонких актиновых протофибриллей. Механизм сокращения сердечной мышцы такой же, как и скелетной, хоть они и различны по своему происхождению в фило- и онтогенезе. Клетки очень богаты митохондриями, обеспечивающими их активную работу. Как и в скелетных мышечных волокнах, здесь имеется вокруг волокон сарколемма, но выражена она слабее. Между волокнами находится очень нежная соединительная ткань, содержащая многочисленные лимфатические сосуды.
У беспозвоночных и позвоночных животных встречается мышечные элементы эпителиального происхождения. Например, у гидры (кишечнополостного животного) тело состоит из двух листков пограничной ткани: наружного - эктодермы и внутреннего - энтодермы. В указанных пластах залегают так называемые эпителиально-мышечные клетки. Их ядросодержащие части подобны клеткам обычного низкоцилиндрического эпителия, а базальные расширены в широкую подошву. В цитоплазме этой подошвы, по существу саркоплазме, проходят многочисленные миофибриллы. Клетки эти, соприкасаясь друг с другом, с одной стороны, образуют настоящий пласт пограничной ткани, а с другой - их подошвы формируют слой особого сократительного аппарата, за счет которого и осуществляются движения тела этого животного.
У позвоночных животных, например, у амфибий в кожных железах имеется два слоя клеток. Внутренний слой представлен типичными сецернирующими клетками, вырабатывающими слизистый или белковый секрет. Их охватывает второй слой клеток (миоэпителиоцитов), мало отличающихся от обычных гладкомышечных. Как и последние, они обладают типичной веретенообразной формой, их заостренные концы вставлены между такими же концами других клеток, а саркоплазма пронизана гладкими миофибриллами. Функция этих миоэпителиальных клеток - выжимание секрета, накапливающегося в полости альвеол в результате деятельности внутреннего слоя клеток. На эпителиальное происхождение этих клеточных элементов указывает, в частности, месторасположение базальной мембраны, которая всегда образуется на границе пограничной ткани и ее физиологической подстилки - рыхлой соединительной ткани. В простых альвеолярных кожных железах земноводных базальная мембрана обнаруживается не между секретирующими и миоэпителиальными клетками, а на границе миоэпителиальных клеток и окружающей рыхлой соединительной ткани.
Наконец, в группу мышечных тканей входят тканевые образования и нервного происхождения - мионевральные клеточные элементы. Всем хорошо известно, что зрачок глаза - отверстие в радужине - меняет свой диаметр в зависимости от интенсивности освещения. Обусловлено это сокращением мышечных клеток, залегающих в толще радужной оболочки. Одни из них расширители, дилятаторы, лежат радиально, другие - суживатели, сфинктеры, располагаются кольцеобразно. В их цитоплазме имеются миофибриллы, вызывающие сокращение этих клеток, в функциональном отношении чисто мышечных. Интересно, что и в этом случае характер миофибриллей связан с быстротой сокращения мышечных элементов. У млекопитающих мионевральные клетки радужины сокращаются медленно и миофибриллы в них гладкие. У птиц сокращение тих клеток происходит быстро и миофибриллы в их мионевральных элементах поперечнополосатые. Возьмем для примера сокола. С большой высоты он пикирует на летящую добычу, не теряя ее из виду. В период этого быстрого падения, когда в несколько секунд хищник проходит полукилометровую дистанцию, резко меняется освещенность фиксируемой его глазом картины. Быстрота сокращения миофибрилл мионевральных элементов соколиного глаза обеспечивает необходимую степень освещенности объекта преследования.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'