Глава X. Ткани внутренней среды организма
Ткани внутренней среды, или опорно-трофические, так же древни по своему происхождению, как и пограничные. Филогенетически они развились из фагоцитобласта Паренхимеллы И. И. Мечникова и по самым характерным чертам своей структуры противоположны эпителиям.
В предыдущей главе мы показали, что пограничные ткани представляют собой оплошные клеточные пласты, расположенные на границе внутренней среды организма и окружающей его среды. Они являются барьерами, через которые осуществляется непрерывный обмен между организмом и наружной средой его обитания. Поэтому клетки в пластах и сами пласты обладают отчетливо выраженной физиологической и связанной с ней морфологической полярностью. Ткани внутренней среды, наоборот, характеризуются сильным развитием межклеточного вещества, представленного волокнистыми структурами (коллагеновыми, эластическими и ретикулярными волокнами) и "аморфной" субстанцией. Классифицируются они по особенностям межклеточного вещества, обусловленным функциональным значением этих тканей. В трофических тканях внутренней среды оно жидкое (кровь) или рыхловолокнистое (ретикулярная и рыхлая соединительная ткань). В механических тканях внутренней среды оно плотноволокнистое (сухожилие, дерма, связка) или богато мукополисахаридами, придающими ему большую плотность (хрящевая ткань), либо, будучи волокнистым, дополнительно обызвествлено (костная ткань).
В отличие от пограничных ткани внутренней среды в своем строении аполярны.
Наконец, система тканей внутренней среды едина в своем происхождении - все они развиваются в эмбриогенезе из мезенхимы, в то время как в группу пограничных тканей входят эпителии разных типов, разной тканевой природы, развивающихся из разных эмбриональных закладок.
Само название возникающей из разных отделов мезодермы эмбриональной мезенхимы в переводе с греческого означает "заполняющая середину". Эмбриональная мезенхима - это как бы первичная соединительная ткань зародыша, заполняющая все промежутки между его плотными зачатками. Она представлена длинными вытянутыми клетками веретенообразной или многоотростчатой формы. Эти клетки залегают рыхло, перекрещиваясь в различных направлениях и спаиваясь своими концами (рис. 45А). Они образуют в совокупности более или менее густую трехмерную сеть. В результате очень ранней дифференцировки (рис. 45Б) эти первоначально одинаковые клетки разделяются на четыре вида (клеточных элементов: 1) округлые, подвижные, передвигающиеся с помощью псевдоподий (ложноножек), подобно амебам - первичные блуждающие клетки; 2) все более уплощающиеся клетки, образующие благодаря стыку друг с другом тонкие однослойные стенки первичных кровеносных сосудов зародыша - первичный эндотелий; 3) оказавшиеся внутри формирующегося кровеносного сосуда и округлившиеся первичные кровяные клетки; 4) оседлые клетки молодой развивающейся соединительной ткани эмбриона. Все эти клетки дифференцированы очень лабильно и могут легко превращаться друг в друга, а первичные блуждающие клетки, проникнув в сосуд, становятся первичными кровяными клетками и наоборот. Первые кровеносные сосуды имеют колбасовидную форму, они сравнительно коротки и слепы на обоих концах. Разрастаясь, строящиеся сосуды приходят своими концами в контакт друг с другом. В дальнейшем перегородки между концами соседних сосудов разрушаются и в результате возникает сеть, по которой начинает циркулировать кровь. Под влиянием работы развивающегося к этому времени сердца сосуды постепенно приобретают форму цилиндрических трубочек. Сначала все сосуды, а также и сердце образованы одним эндотелием. Затем стенки их утолщаются и усложняются путем образования вокруг эндотелиальной трубки за счет окружающей мезенхимы разно построенных оболочек, структура которых строго отвечает местным условиям кровотока.
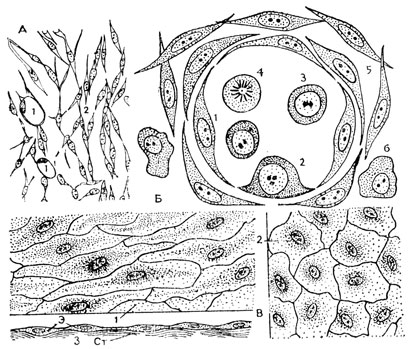
Рис. 45. А - эмбриональная мезенхима (2), в ней новообразованные капилляры (1). Б - дифференцировка мезенхимы: 1 - мезенхимная клетка, становящаяся эндотелиальной; 2 - превращение эндотелиальной клетки в первичную кровяную клетку (3): 4 - митоз первичной кровяной клетки; 5 - мезенхимные клетки, 6 - первичная блуждающая клетка. В - эндотелиальная выстилка разных крупных сосудов: 1 и 2 - вид с поверхности; 3 - вертикальное сечение эндотелиального пласта; Э - эндотелиальные клетки; СТ - подлежащая соединительная ткань
Сосудистая система обеспечивает движение крови по телу, регулирует ее поступление в разные органы и осуществляет обмен веществ между последними и кровью. В этой системе имеются центральный орган (сердце), приводящий кровь в движение, кровеносные сосуды (артерии, несущие кровь от сердца, и вены, по которым она возвращается в сердце) и важнейшая ее часть - кровеносные капилляры, выполняющие трофическую (питающую) функцию. Входящие в состав этой же системы лимфатические сосуды и капилляры являются только приводящими. Они выполняют роль дренажного аппарата, компенсирующего работу кровеносной системы. Плазма крови, просачивающаяся через стенки кровеносных капилляров в ткани и становящаяся там тканевой лимфой, насасывается затем в лимфатические капилляры и уже в виде сосудистой лимфы возвращается обратно в кровоток, изливаясь через грудной лимфатический проток в верхнюю полую вену.
В сформировавшейся сосудистой системе особое место принадлежит эндотелию, из которого построены кровеносные и лимфатические капилляры и который образует граничащую с кровью или лимфой внутреннюю выстилку всех кровеносных и лимфатических сосудов и сердца.
Эндотелий, выполняющий во внутренней среде организма ограничительную функцию, приобрел конвергентно большое сходство с мезотелием, то есть построен по типу однослойного плоского эпителия (рис. 45В). Образовавшись в процессе эмбрионального развития из мезенхимы, дифференцированный эндотелий ведет себя в дальнейшем как самостоятельная ткань, способная к разрастанию только за счет размножения своих собственных клеток. Последнее обстоятельство дало основание некоторым исследователям относить эндотелий к пограничным тканям, рассматривая его как эпителий особого типа, погруженный во внутреннюю среду организма.
Эндотелиальные клетки (эндотелиоциты), образующие стенки кровеносных капилляров, в одних органах имеют многочисленные микроворсинки и проявляют активный пиноцитоз, что свидетельствует об их высокой всасывающей способности. В других случаях для эндотелия характерно наличие пор, локализующихся преимущественно в периферических участках цитоплазмы эндотелиальных клеток. Количество их может сильно варьировать и в некоторых органах (в почке, гипофизе) на 1 кв. мк поверхности клетки находится до 12 таких пор, что чрезвычайно повышает проницаемость эндотелиальной стенки. Под эндотелиальными клетками имеется базальная мембрана, что еще больше увеличивает сходство данной ткани с мезотелием.
Эндотелий лимфатических капилляров, представляющих собой слепо начинающиеся мешки, обеспечивает их дренажную функцию, ибо насасывание тканевой лимфы внутрь лимфатических капилляров происходит в результате активной деятельности эндотелия.
Эндотелий, выстилающий просвет артерий, выполняет и здесь трофическую функцию - вся стенка более мелких артерий и внутренняя половина или треть стенки "рунных сосудов питаются с помощью эндотелия за счет протекающей крови.
В кроветворных органах (костном мозге и в еще большей степени в селезенке и лимфатических узлах), где протекающие через них жидкости не только обогащаются кровяными клетками, но и очищаются от посторонних частичек, в том числе микробов, входящий в их состав эндотелий обладает способностью к фагоцитозу.
Кровь - это жидкость, на которую приходится 1/2 общего веса тела. Она почти наполовину состоит из клеток, а другую половину составляет кровяная плазма. В кровяной плазме, а также в сосудистой и тканевой лимфе содержатся белок, соли и различные продукты обмена веществ. Из солей главное значение имеет хлористый натрий, или поваренная соль. Общее осмотическое давление в крови млекопитающих соответствует давлению, равному девяти десятым процента (раствора поваренной соли. Такой раствор называют поэтому физиологическим и возмещают им кровопотери. В крови встречается сахар, разносимый ею по всему организму из его депо (источником его является животный крахмал - гликоген, залегающий в соматических мышцах и печени) или из кишечника, а также такие продукты обмена, как мочевина, холестерин, лецитин и др. Кровью разносятся ко всем клеткам тела витамины - вещества, необходимые для их нормальной жизнедеятельности, и гормоны. В кровяной плазме содержится в растворе белок фибриноген. Под влиянием специального фермента тромбокиназы, образуемой распадающимися тромбоцитами (особыми форменными элементами крови), фибриноген в присутствии солей кальция выпадает в осадок в виде тонких нитей белка фибрина. Такой процесс приводит к свертыванию крови, т. е. образованию пробки, которая приостанавливает кровотечение, возникающее при ранении. Кровяная плазма, лишенная фибриногена, называется кровяной сывороткой. Аналогичным образом сворачивается и лимфа. Если травма кожи носит поверхностный характер и кровеносные сосуды при этом не задеты, на поверхность пропотевает прозрачная желтоватая жидкость. Это и есть тканевая жидкость, или тканевая лимфа. Спустя какое-то время она тоже свертывается и закрывает таким образом поврежденный участок.
Форменные элементы крови млекопитающих и человека представлены эритроцитами, то есть красными кровяными клетками, лейкоцитами, или белыми кровяными клетками, и кровяными пластинками, обозначаемыми как тромбоциты по аналогии с тромбоцитами крови низших позвоночных.
Из форменных элементов крови громадное большинство составляют красные кровяные тельца. В норме у человека на 1 мм3 крови их приходится около 5 млн. у мужчин и 4,5 млн. у женщин. У млекопитающих и человека они представляют собой мелкие круглые двояковогнутые пластинки или неглубокие колпачки. Размер их у человека 7,3 мк. Самые мелкие эритроциты у козы, их диаметр всего 4 мк, зато количество красных кровяных клеток в 1 мм3 составляет 14,5 млн. У млекопитающих и человека эритроциты лишены ядер. Срединная часть их тоньше, чем периферия, и поэтому кажется светлее.
Эритроциты - это переносчики кислорода. Их тело заполнено особым веществом: пигментом белковой природы - гемоглобином. Гемоглобин легко вступает в соединение с кислородом и легко его отдает. Гемоглобин, присоединивший к себе кислород, называют оксигемоглобином. В легких пузырьки (альвеолы), сидящие на мельчайших разветвлениях бронхиального дерева, являются местом, где происходит газообмен. Стенки пузырьков густо оплетены кровеносными капиллярами. Здесь кислород из воздуха, вдохнутого легкими, через тонкую эпителиальную выстилку альвеолы и эндотелий капилляра захватывается гемоглобином эритроцитов, проходящих по капилляру. Гемоглобин, становясь оксигемоглобином, принимает ярко-красный цвет и придает крови алую окраску. Далее кислород разносится эритроцитами по всему телу и им снабжается каждая клетка. Будучи непрочно связанным с гемоглобином, он легко переходит в клетки и ткани, которые жадно его поглощают.
В капиллярах происходит не только отдача кровью кислорода, но и поглощение углекислоты и других продуктов клеточного обмена из тканей. Углекислота в основном соединяется с кровяной плазмой и лишь частично с эритроцитами. В легком эта углекислота из капилляров поступает в альвеолы и вместе с водяными парами удаляется из организма ори выдохе.
У всех позвоночных животных, кроме млекопитающих, эритроциты являются настоящими клетками и содержат ядра, поэтому сбоку выглядят двояковыпуклыми. Форма их не округлая, а эллипсоидная. Особенно крупных размеров эритроциты достигают у земноводных. У лягушки, например, их больший диаметр равен 23 мк, а меньший - 16 мк. Понятно, что функция газообмена крупными эритроцитами выполняется гораздо менее совершенно, чем мелкими, так как при большом размельчении какого-либо тела отношение его поверхности к объему резко возрастает. А захват или отдача кислорода эритроцитом происходит через его поверхность. В этом отношении более приспособленными к своей функции являются эритроциты млекопитающих. К тому же их тело представляет собой по существу мешочек, целиком заполненный гемоглобином. Интересно отметить, что у некоторых млекопитающих (верблюдов) красные кровяные "летки безъядерные, но имеют эллипсоидную форму.
Таким образом, эритроциты являются высоко дифференцированными клетками, то своему строению наиболее отвечающими выполняемой ими функции разносчиков кислорода. Лишенные ядра с цитоплазмой, почти полностью загруженной гемоглобином, эритроциты все же довольно долго выполняют свою функцию, переживая около 100 дней в кровяном русле. Отмершим приходят на смену новые красные кровяные клетки, образующиеся в костном мозге. Благодаря интенсивно протекающему процессу кроветворения в крови поддерживается постоянное количество эритроцитов, в среднем отвечающее норме.
Лейкоциты представляют собой сборную группу кровяных клеток. Число их в крови примерно в тысячу раз меньше, чем эритроцитов. В норме оно колеблется от 6 до 8 тыс. в 1 мм3. Большую часть, около 3/4, составляют так называемые сегментоядерные формы, т. е. клетки с ядром, состоящим из фрагментов, соединенных перемычками. Раньше такие клетки ошибочно рассматривали как многоядерные и называли полинуклеарами. Сегментоядерные лейкоциты называются еще зернистыми лейкоцитами, или гранулоцитами, так как в их цитоплазме находятся многочисленные зернышки, гранулы. Содержимое этих гранул представляет собой особые вещества ферментативного характера. При гибели лейкоцита они высвобождаются из его тела и начинают проявлять свое лизирующее, растворяющее действие. Гранулы у разных лейкоцитов различные и, в частности, проявляют разную склонность к кислым и основным красителям. Соответственно этому гранулоциты делят на 3 вида: нейтрофилы, ацидофилы (эозинофилы) и базофилы. Все лейкоциты, находясь в жидкости, обладают шарообразной формой, а на мазках уплощены.
Нейтрофильные гранулоциты (нейтрофилоциты) имеют на мазках диаметр 10 - 12 мк, обладают пылевидной зернистостью. Их гранулы слабо красятся как кислыми, так и основными красками. Размер гранул 0,2 - 0,5 мк. Ядра у неитрофилов наиболее сегментированы, причем чем старше клетка, тем из большего количества сегментов состоит ее ядро. Число их может доходить до 5. У молодых же неитрофилов ядра похожи на изогнутую палочку, еще не сегментированную, поэтому такие клетки называют палочкоядерными. В цитоплазме неитрофилов с помощью электронного микроскопа обнаруживаются различные органоиды. Имеется значительное количество мелких митохондриев и пузырьков слабо развитого эндоплазматического ретикулума. Некоторые из гранул представлены лизосомами. Нейтрофилы являются активнейшими фагоцитами. При попадании инфекции во внутреннюю среду тела они первыми устремляются к месту внедрения инородного, покидая сосудистое русло. Если это мелкие образования, например, бактерии, нейтрофилы захватывают их и переваривают. Если это более крупное тело, например, деревянная заноза, нейтрофилы окружают его более или менее мощным слоем, создавая защитный лейкоцитарный вал. Часть их, в особенности нейтрофилы, прилежащие к инородному телу, погибают и рассыпаются, выбрасывая таким путем наружу свои переваривающие, лизирующие ферменты.
Реакция организма, направленная на уничтожение вредности, получила наименование воспалительной. Первая клеточная воспалительная реакция организма - это нейтрофильная. Вот почему они получили еще 2 названия - специальных лейкоцитов (специальная функция - фагоцитарная) и микрофагов. Последнее название принадлежит великому русскому ученому И. И. Мечникову, который в 1883 высказал идею о роли фагоцитов в организме многоклеточных животных. Он показал, что присущая одноклеточным формам способность поддерживать свое существование путем внутриклеточного пищеварения в многоклеточном организме переключилась на защитную фагоцитарную функцию. Питание такие клетки-защитники получают от организма, частью которого они являются. Способность их захватывать и переваривать микробы теперь служит интересам целого, его освобождению, очистке от инфекции. Нейтрофилы - самые распространенные из лейкоцитов. Количество их у человека колеблется в норме от 63 до 67%, то есть составляет приблизительно 2/3 от общего количества белых кровяных клеток.
Процесс прохождения лейкоцитов через стенку кровеносного сосуда получил наименование диапедеза. При возникновении очага воспаления в прилежащих к нему кровеносных сосудах лейкоциты замедляют свою циркуляцию и начинают прилипать к сосудистой стенке на стороне, обращенной к очагу воспаления, от которого исходят какие-то химические вещества. Это явление обозначают как пристеночное стояние лейкоцитов. Затем они начинают как бы вбуравливаться в толщу стенки кровеносного сосуда и пробираются через ее структурные элементы наружу в соединительную ткань.
Следующими по количеству из гранулоцитов являются ацидофильные лейкоциты, или эозинофилы. Их в норме 3 - 4%. Число их резко возрастает при заражении организма глистами. Клетки эти содержат обычно двухсегментное ядро, те же органоиды, что и нейтрофилы, но переполнены крупными округлыми блестящими гранулами размером 0,7 - 1,3 мк. Последних так много, что эозинофилы плохо фагоцитируют и оказывают свое защитное действие преимущественно распадом и освобождением из цитоплазмы зернышек, содержащих ферменты, с помощью которых они могут обезвреживать токсины. По той же причине эозинофилы являются менее подвижными клетками, чем нейтрофилы. Размеры их несколько больше, чем у нейтрофилов, и составляют на мазках 12 - 14 мк в диаметре.
Меньше всего в крови базофилов - 0,5 - 1%. Они обладают и наименее сегментированным ядром, которое по своей изогнутой форме напоминает латинскую букву S. Крупные гранулы базофилов размером 0,8 - 1 мк красятся основными красками (употребляются для этого синие красители, например, азур) в отличающийся от них тон (фиолетовый) - свойство, получившее наименование метахромазии. В гранулах установили наличие гистамина и гепарина. Присутствие в базофилах гепарина говорит о влиянии их на свертывание крови. Размер этих клеток почти такой же, как у нейтрофилов, - диаметр на мазках 11 - 12 мк.
Вторую большую группу лейкоцитов составляют незернистые формы, или агранулоциты. Ядра их не сегментированы, а гранулы в цитоплазме отсутствуют. Основную массу составляют лимфоциты, число которых у взрослых людей варьирует от 24 до 30%. В организме встречаются 3 разновидности лимфоцитов - большие, средние и малые. В нормальной крови большие лимфоциты (их размер превышает 10 мк) не циркулируют, они залегают лишь в органах лимфоидного кроветворения. Малые и средние лимфоциты отличаются друг от друга морфологически главным образом количеством цитоплазмы. В малых лимфоцитах она представлена тонким ободком цитоплазмы, окружающим округлое и компактное ядро, богатое глыбками хроматина. Эти клетки имеют в диаметре 5 - 6 мк. В средних лимфоцитах цитоплазмы больше, ядро светлее и лежит несколько эксцентрично (рис. 30-7). Их размер 7 - 10 мк. В лимфоцитах электронномикроскопически обнаруживаются все клеточные органоиды, хотя они в общем слабо развиты.
Лимфоциты выполняют в организме защитную роль, а также являются стимуляторами тканевых процессов при посттравматической регенерации. Обе эти функции осуществляются ими не в кровяном русле, а в окружающей кровеносные сосуды рыхлой соединительной ткани. Они менее подвижны, чем нейтрофилы, и образуют вторую волну блуждающих клеток крови, продвигающихся к инородному телу, проникшему во внутреннюю среду организма. Если специальные лейкоциты оказываются в формирующемся очаге воспаления в первые же часы после попадания инфекции, то лимфоцитам на это требуется несколько часов. Это объясняется тем, что лимфоциты, особенно малые формы, бедны цитоплазмой, псевдоподии их невелики, и поэтому передвижение данных клеток по направлению к источнику раздражения замедленное. Следует отметить, что по мере приближения к очагу воспаления лимфоциты постепенно увеличиваются в размерах, особенно нарастает в них количество цитоплазмы. Они превращаются в крупных фагоцитов, достигающих 20 мк и названных поэтому И. И. Мечниковым макрофагами.
Лимфоциты способны превращаться и в другие клеточные формы соединительной ткани и даже в клетки гладкой мышечной ткани, которая, как и соединительная, имеет мезенхимное происхождение. По-видимому, к такого рода перестройкам способны не все лимфоциты, а лишь малодифференцированные элементы, играющие роль камбия, внешне не отличимые от клеток, заканчивающих свой жизненный путь. Считают, что камбиальные элементы в основном имеют вид малых лимфоцитов.
Выполняя другую свою функцию активаторов тканевых процессов, лимфоциты являются источниками особых веществ - трефонов. Под влиянием этих трефонов, как показал Г. К. Хрущев, ускоряются процессы заживления ран. В ранах происходит накопление лимфоцитов, ткани инфильтрируются ими и поступающие в окружающую среду из этих клеток трефоны стимулируют репаративные процессы в регенерирующих тканях.
Последняя клеточная форма из числа агранулоцитов - моноциты (6 - 8% от общего числа лейкоцитов). Они являются наиболее крупными из лейкоцитов: на мазках диаметр может достигать 20 мк. Ядра моноцитов бобовидной формы и лежат эксцентрично. Цитоплазмы в них много. При исследовании с помощью электронного микроскопа в ней обнаруживаются различные органоиды. Под световым микроскопом особенно хорошо заметна центросома, которая оттесняет ядро из срединного положения в клетке. В цитоплазме встречается так называемая азурофильная зернистость, ибо гранулы этих клеток обнаруживают склонность к основной краске - азуру. Прошедшие через стенку кровеносного сосуда моноциты по своему виду ничем не отличаются от макрофагов и обладают всеми присущими этой клеточной форме свойствами, но они могут фагоцитировать и в самом кровяном русле. Таким образом, моноциты можно рассматривать как макрофаги крови.
Указанное выше процентное содержание разных форм лейкоцитов в крови характеризует состояние крови у здорового человека. При разного рода заболеваниях наблюдается количественный сдвиг тех или иных из них в ту или другую сторону, что служит одним из важных диагностических признаков. Приводим так называемую лейкоцитарную формулу здорового человека, утвержденную Министерством здравоохранения СССР: базофилы 0 - 0,5%; эозинофилы 3 - 4%; нейтрофилы палочкоядерные 4% и сегментоядерные 63 - 67%; лимфоциты 24 - 30%, моноциты 6 - 8%.
Третьей группой форменных элементов крови являются тромбоциты, которые у человека представляют собой не клетки, а как бы клеточные обломки, и поэтому обозначаются как кровяные пластинки. Функционально же кровяные пластинки млекопитающих и человека однозначны тромбоцитам всех других классов позвоночных. Они обладают свойством слипания в массы, образуя тромбы, а главное, дают начало ферменту тромбокиназе, который участвует в процессе свертывания крови. У человека кровяные пластинки имеют очень мелкие размеры - 2 - 3 мк, зато число их велико. Колеблется оно в больших пределах - от 150000 до 300 000 в 1 мм3.
Тромбоцит имеет вид отростчатого обрывка, содержащего внутри некоторое количество зернышек. Эту срединную часть тромбоцита называют грануломер, периферическая гомогенная часть за ее стекловидный характер получила наименование гиаломера. В грануломере обнаруживаются электронномикроскопически митохондрии и немногочисленные элементы эндоплазматической сети. В настоящее время принято различать среди тромбоцитов млекопитающих и человека разные формы - юные, зрелые, старые, а также гигантские формы раздражения и дегенеративные. У юных форм грануломер относительно невелик, у старых, наоборот, содержит большое количество зерен.
В настоящее время многочисленными исследованиями, проведенными в клинике, и экспериментами на животных показано, что тромбоцитарная формула - тромбоцитограмма, то есть процентное соотношение разных форм кровяных пластинок, может иметь определенное значение для ранней диагностики злокачественных новообразований. Особенно наглядно это видно в экспериментах с перевивными злокачественными опухолями. Уже вскоре после перевивки процент старых форм резко повышается. Если в норме их обычно бывает не более 4 - 5%, то при наличии в организме злокачественных разрастаний процент старых форм может достигать многих десятков. Критической точкой считается 7% их содержания от общего числа кровяных пластинок. Количество старых форм тромбоцитов, превышающее 7%, должно настораживать, и человека с таким их содержанием необходимо всесторонне обследовать на рак и держать постоянно под контролем врача. Интересно, что после удаления у экспериментального животного перевивной злокачественной опухоли в крови устанавливается нормальное соотношение разных форм тромбоцитов, а процент старых ниже 7. То же самое наблюдается и в клинике после успешного и полного удаления раковой опухоли у больных людей.
Образование форменных элементов крови у взрослых людей происходит в специальных кроветворных органах. Одни из них - костный мозг и селезенка - лежат на пути кровеносного русла, другие - лимфатические узлы - расположены по ходу лимфатических сосудов. В костном мозге протекают процессы новообразования эритроцитов (эритропоэз), гранулоцитов (гранулоцитопоэз) и кровяных пластинок (тромбоцитопоэз). Кроветворную ткань костного мозга называют миелоидной (по-гречески "миелос" означает костный мозг), а костномозговое кроветворение - миелоидным. Лимфоциты образуются в селезенке и лимфатических узлах. Кроветворную ткань этих органов обозначают как лимфоидную, а кроветворение в ней (лимфоцитопоэз) лимфоидным. Моноциты могут возникать в миелоидной и в лимфоидной кроветворных тканях.
Согласно наиболее распространенной теории кроветворения миелоидная и лимфоидная ткани имеют общие родоначальные клетки, детерминированные как кроветворные. Эти, как их называют, стволовые клетки по своему виду напоминают малые лимфоциты. Установлено, что широкопотенциальные S-клетки постоянно выселяются из разных участков костного мозга и мигрируют по организму, частично попадая в кровь. Так происходит непрерывно обмен стволовыми клетками разных участков кроветворной системы и заселение ими лимфоидной ткани.
Важнейшей особенностью стволовых клеток является их свойство определять величину своего пула (то есть своей совокупности в организме), способного к неограниченному размножению, и регулировать пролиферацию (размножение клеток) в соответствии с потребностями организма. Другая, столь же важная особенность стволовых клеток заключается в их способности дифференцироваться в полустволовые клетки эритроидного, гранулоцитарного, мегакариобластического (мегакариоциты - гигантские клетки костного мозга, с которыми связан тромбоцитопоэз) и лимфоидного направления. Эти полустволовые клетки, в отличие от стволовых, обладают узкой кроветворной детерминацией и обнаруживают ограниченную способность к самоподдержанию. Они постоянно пополняются за счет S-клеток. Выбор направления дифференцировки S-клеток в значительной мере определяется структурой данной клеточной популяции. Решающая роль принадлежит механизму обратной связи - при достаточном количестве форменных элементов крови какого-то ряда дифференцировка S-клеток в сторону образования полустволовых клеток с подобной детерминацией тормозится. Наоборот, при нехватке каких-либо клеток крови дифференцировка соответствущих полустволовых клеток из стволовых резко усиливается. Однако часть S-клеток, пролиферируя в зависимости от потребностей организма с меньшей или большей интенсивностью, на протяжении всей жизни особи сохраняет свой стволовой характер.
Полустволовыми клетками являются гемоцитобласты, которыми начинаются ряды эритропоэза, гранулоцитопоэза, лимфоцитопоэза и которые дают начало мегакариоцитам. Гемоцитобласты лимфоидной ткани называют лимфобластами. По строению они весьма сходны с гемоцитобластами костного мозга, но отличаются от них потенциями развития.
Гемоцитобласты достигают по своим размерам 15 мк. Они отличаются слабобазофильной цитоплазмой и наличием крупного более или менее округлого ядра. Цитоплазмы относительно немного, но электронномикроскопически в ней обнаруживаются все органоиды. Гемоцитобласты способны к амебоидному движению.
Процесс дифференцировки гемоцитобластов в любом направлении связан с целой цепью последовательных митозов, причем после каждого митоза дочерние клетки оказываются более дифференцированными, чем материнская.
При эритропоэзе (рис. 46) клетки, возникшие путем деления гемоцитобластов, отличаются неспособностью к амебоидному движению, а главное - резкой базофилией цитоплазмы (в результате усиленного образования рибосом, как подготовки к синтезу белка - гемоглобина). Ядро у них округло. Эти клетки называют проэритробластами, или базофильными эритробластами. В результате повторного размножения их и нарастания количества гемоглобина появляется ряд эритробластов, именуемых полихроматофильными. При общепринятой окраске крови эозин-азуром гемоглобин окрашивается эозином в красный цвет. Базофильная цитоплазма, наоборот, окрашивается азуром в синий цвет. По мере увеличения количества гемоглобина в цитоплазме общая окраска эритробластов изменяется от фиолетовой к сиреневой. По размерам эти клетки уступают проэритробластам. Дальнейшее накопление в цитоплазме гемоглобина, обусловливающее оксифильную окраску цитоплазмы, приводит к образованию ацидофильных эритробластов.
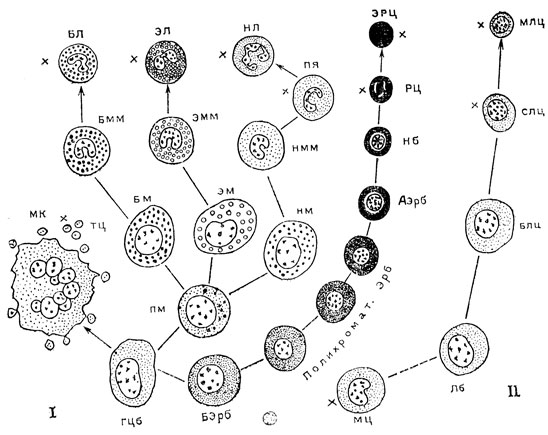
Рис. 46. Схема последовательных стадий кроветворения в красном костном мозге (I) и в лимфоидной ткани (II). ГЦб - гемоцитобласт; МК - мегакариоцит (гигантская клетка костного мозга); ТЦ - тромбоциты (кровяные пластинки); ПМ - промиелоцит; БМ, ЭМ, НМ - базофильный, ацидофильный и нейтрофильный миелоциты; БММ, ЭММ, НММ - базофильный, эозинофильный и нейтрофильный метамиелоциты; ПЯ - палочкоядерный нейтрофил; БЛ - базофил; ЭЛ - эозинофил; НЛ - сегментоядерный нейтрофил; БЭрб - базофильный эритробласт (проэритробласт); Эрб - полихроматофильные эритробласты; АЭрб - ацидофильный эритробласт; Нб - нормобласт; РЦ - ретикулоцит; ЭРЦ - эритроцит; ЛБ - лимфобласт (гемоцитобласт лимфоидной ткани); БЛЦ, СЛЦ, МЛЦ - большой, средний и малый лимфоциты; МЦ - моноцит. Знаком '×' отмечены форменные элементы (клетки), встречающиеся в крови здоровых людей
Одни из ацидофильных эритробластов перестают делиться и задерживаются в костном мозге на этой стадии эритропоэза. Другие делятся дальше и в результате дифференцирующего митоза возникают клетки, близкие по размерам и количеству гемоглобина к эритроцитам, но содержащие ядро, - нормобласты. Молодые нормобласты еще продолжают делиться. Затем ядра в нормобластах уплотняются, пикнотизируются и выбрасываются из клетки. Нормобласт становится молодым эритроцитом (ретикулоцитом) крови. В нем еще обнаруживаются под электронным микроскопом в остатках цитоплазмы немногочисленные митохондрии и элементы эндоплазматического ретикулума. Именно благодаря им в ретикулоцитах крови при специальной окраске появляется зернисто-сетчатая субстанция (гранулоретикулофиламентозное вещество). Наконец, ретикулоцит крови теряет остатки органоидов, сохраняя лишь плазмолемму, и превращается в эритроцит.
При гранулоцитопоэзе гемоцитобласты, делясь митотически, образуют промиелоциты - клетки с овальным ядром, слабобазофильной цитоплазмой, хорошо заметной центросомой и неспецифической зернистостью. В процессе дальнейших дифференцирующих митозов в клетках появляется все в большем количестве специфическая зернистость и они становятся миелоцитами - нейтрофильными, ацидофильными и базофильными. Эти клетки продолжают делиться и дают начало метамиелоцитам, которые характеризуются способностью к активному амебоидному движению и такой степенью дифференцировки, что уже не могут делиться. Нейтрофильные и эозинофильные метамиелоциты обладают подковообразно-колбасовидными ядрами. В процессе дальнейшей дифференцировки они проходят стадию палочкоядерных лейкоцитов. Развитие завершается образованием зрелых сегментоядерных гранулоцитов.
Тромбоцитопоэз связан с образованием из гемоцитобласта мегакариобласта, отличающегося крупными размерами и лапчатым ядром. Резко увеличиваясь в размерах в результате митозов, не завершающихся разделением клеточного тела, мегакариобласт превращается в мегакариоцит - гигантскую клетку с крупными сегментированными ядрами и множеством центриол. Цитоплазма мегакариоцитов образует многочисленные выросты диаметром 2 - 3 мк, которые, отрываясь, становятся кровяными пластинками.
Лимфоцитопоэз начинается с митотических делений гемоцитобластов, в результате которых возникают большие лимфоциты. Митотические деления больших лимфоцитов приводят к образованию средних лимфоцитов, а последние, делясь митотически, дают начало малым лимфоцитам.
До настоящего времени точно не установлены источники образования моноцитов. Считают вероятным их происхождение из гемоцитобластов лимфоидной ткани.
Помимо крови и лимфы трофическую роль в организме выполняют разные виды рыхлой соединительной ткани. Самая нежная из этих тканей носит название ретикулярной соединительной ткани (по-гречески "ретикулум" означает сеть). Она ближе других тканей стоит к эмбриональной мезенхиме и также имеет вид трехмерной сети, т. е. сети, петли которой лежат не в одной плоскости, а в трех измерениях пространства (рис. 47 Б). Основное морфологическое отличие ее от мезенхимы состоит в том, что через клеточные тела, а отчасти между ними проходят ретикулярные волокна. Последние образованы очень тонкими (400 Å) волоконцами, спаянными друг с другом особым веществом. Эти волокна имеют еще и другие наименования, оттеняющие ту или иную их способность: ретикулиновые - по характеру образующего их белка ретикулина; аргирофильные, или аргентофильные, то есть выявляющие склонность к серебру. При пропитывании ткани раствором азотнокислого серебра и последующем восстановлении металла из раствора металлическое серебро выпадает на этих волокнах в виде мелких частиц черного цвета. Благодаря слою черни ретикулярные волокна становятся хорошо заметными на общем коричневом фоне. Напоминаем, что такой способ обнаружения определенных структур называется импрегнацией. Варьированием методики импрегнации можно выявлять и другие гистологические образования, в частности, аппарат Гольджи или границы между клетками.
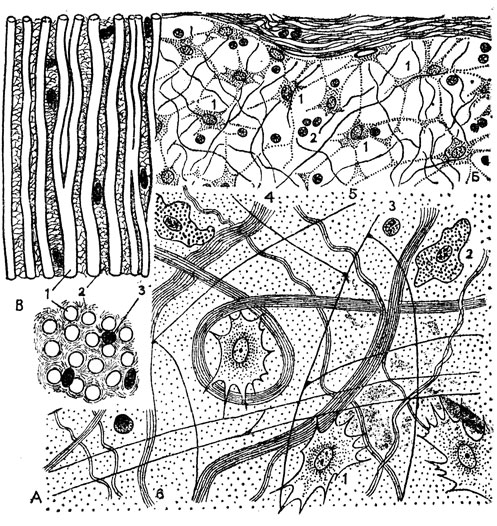
Рис 47. Соединительные ткани. А - рыхлая волокнистая соединительная ткань: 1 - фибробласт; 2 - гистиоцит; 3 - малый лимфоцит; 4 - пучок коллагеновых волокон; 5 - эластическое волокно; 6 - основное 'аморфное' вещество. Б - ретикулярная ткань в лимфатическом узле: 1 - ретикулярные клетки; 2 - лимфоциты. В - продольный и поперечный разрезы плотной волокнистой соединительной ткани эластического типа (на схеме она для наглядности разрыхлена): 1 - эластические волокна; 2 - нежные прослойки рыхлой соединительной ткани между волокнами; 3 - ядра клеток
Ретикулярные волокна образуются внутри ретикулярных клеток. Вне клеток оказываются те их участки, с которых клетки сползли. Дело в том, что ретикулярные клетки обладают способностью высвобождаться из связи друг с другом, сниматься с места и превращаться в блуждающие клеточные элементы, которые не отличаются от моноцитов, покинувших кровеносное русло, макрофагов.
Ретикулярные клетки (ретикулоциты) обладают еще одной особенностью, сближающей их с макрофагами и с эндотелиальными клетками кроветворных органов и печени - они могут фагоцитировать и нефрофагоцитировать, т. е. накоплять коллоидные частички из окружающей их тканевой жидкости, освобождая внутреннюю среду организма от посторонних элементов. Ретикулярные клетки напоминают мезенхимные и своей способностью давать начало самым различным клеткам тканей внутренней среды.
Раньше ретикулярную соединительную ткань приводили как пример так называемого синцития, соклетия. Считалось, что отростки этих неправильно звездчатых клеток непрерывно переходят друг в друга. С помощью электронного микроскопа было установлено, что между соприкасающимися друг с другом клеточными отростками имеются лишь тесные контакты. Клеточные мембраны здесь не прерываются, а образуют многочисленные взаимные впячивания-выпячивания в виде описанных нами ранее "замков", более или менее прочно удерживающие клетки друг с другом в этой трехмерной сети.
Ретикулярная соединительная ткань составляет основу кроветворных органов. Через нее, например, в лимфатических узлах непрерывно протекает сосудистая лимфа и благодаря этому очищается. Сеть, образуемая ретикулярными клетками, не является просто механическим фильтром, а активно изымает из медленно текущей здесь лимфы как микроскопические, так и коллоидные посторонние частички.
Типичная рыхлая волокнистая соединительная ткань (рис. 47А) имеет большое распространение в организме. Она является, во-первых, как бы упаковочным материалом для элементов других тканей, а во-вторых, выполняет важную трофическую функцию, так как через нее непрерывно протекает тканевая жидкость, несущая клеткам необходимые для их жизни и работы вещества и уносящая продукты их обмена. В соответствии с такой функцией в межклеточном веществе этой ткани волокнистые элементы лежат рыхло и беспорядочно, как в вате и других упаковочных материалах.
Большая часть волокон образована из белка коллагена. Коллагеновые волокна получили свое название за свойство развариваться в клей. Это очень прочные и мало растяжимые волокна толщиной 1 - 3 мк, представляющие собой пучки волоконец, фибрилл, спаянных вместе интерфибриллярным веществом. Электронный микроскоп показал, что сами фибриллы являются пучками тончайших (до 100 Å) протофибрилл. Причем оказалось, что они имеют отчетливую периодичность строения, обнаруживая на электроннограммах поперечную полосатость с периодом исчерченности в 640 Å.
Ни протофибриллы, ни фибриллы, видимые в оптический микроскоп, не ветвятся. Однако пучки коллагеновых волокон могут по ходу своему менять диаметр за счет того, что часть фибрилл в виде пучка низшего порядка покидает основной пучок, причем может присоединиться к другому коллагеновому пучку и пойти с ним в совершенно ином направлении.
Другие волокнистые структуры рыхлой соединительной ткани - эластические волокна обладают свойствами, диаметрально противоположными коллагеновым. Они очень растяжимы и отличаются малой прочностью на разрыв. Свое название они получили за присущее им физическое качество - эластичность. Если коллагеновый пучок может быть в какой-то мере уподоблен канату, эластические волокна напоминают собой резиновые тяжи. Эти волокна под световым микроскопом кажутся гомогенными, но под электронным микроскопом выявляется их протофибриллярное строение. Толщина протофибрилл составляет всего 30 - 40 Å. В отличие от коллагеновых эластические волокна ветвятся. Образующий их белок-эластин в клей не разваривается. Он вообще очень устойчив и у человека не поддается действию пищеварительных ферментов. Эластические волокна в рыхлой соединительной ткани образуют редко-петлистую сеть. В совокупности они создают эластичный каркас, который возвращает ткань в исходное состояние после ее деформаций, связанных, в частности, с движением тела.
В рыхлой волокнистой соединительной ткани, в особенности по ходу сосудов, встречается и небольшое количество ретикулярных волокон, образуемых ретикулярными клетками. Следует отметить, что ретикулярные волокна по составу их белка ретикулина близки коллагеновым и, как последние, обнаруживают под электронным микроскопом поперечную периодическую исчерченность. С другой стороны, они, подобно эластическим волокнам, не перевариваются пищеварительным соком - трипсином и проявляют устойчивость к действию слабых кислот и щелочей. Они более растворимы, чем коллагеновые волокна, но меньше, чем эластические.
Основное межклеточное вещество соединительной ткани не является аморфным, как представляли себе раньше. Электронномикроскопическое исследование показало, что оно не гомогенно, а содержит многочисленные очень мелкие пучки протофибрилл, переплетающихся друг с другом в разных направлениях и залитых в матрикс (от греческого слова "матер" - мать; здесь нужно понимать как маточную среду), образованный, по данным гистохимических исследований, преимущественно из мукоидов. Мукоиды относятся к сложным белкам (протеидам), точнее к мукопротеидам, т. е. таким соединениям, в которых белок связан с углеводами, в данном случае со сложными углеводами - полисахаридами, или более точно мукополисахаридами, в первую очередь с гиалуроновой и хондроитинсерной кислотами.
Вдоль капилляров и мелких кровеносных сосудов располагаются другие камбиальные клеточные элементы рыхлой соединительной ткани - адвентициальные клетки. Из них, главным образом, и возникают фибробласты (клетки-волокнообразователи) соединительной ткани. Они способны дифференцироваться даже в гладкие мышечные клетки.
Рыхлая волокнистая соединительная ткань вообще богата клетками. Их можно разбить на три категории - оседлые, временно оседлые и блуждающие. Ретикулярные (ретикулоциты) и адвентициальные (перициты) клетки должны быть отнесены к числу оседлых, однако основную часть последних и вообще наиболее многочисленную группу клеток рыхлой волокнистой соединительной ткани представляют фибробласты и их более зрелые и дифференцированные формы - фиброциты. Молодые фибробласты близки камбиальным элементам, наиболее дифференцированные заканчивают свой жизненный путь и уже не способны ни к дедифференцировке, ни к делению. По своей функции фибробласты являются клетками, образующими межклеточное вещество соединительной ткани, всех ее волокон и аморфной субстанции, в которую оказываются как бы замурованными и клетки, и волокнистые структуры этой ткани.
Фибробласты - крупные (порядка 20 мк), уплощенные многоотростчатые клетки (рис. 47А), обладающие светлым ядром с пылевидно рассыпанным по нему хроматином. Форма их на разрезе веретенообразная с расширением в месте залегания ядра. Цитоплазма содержит все свойственные клеткам органоиды. Участвуя в образовании основного межклеточного вещества соединительной ткани, фибробласты синтезируют и выделяют составляющие его мукополисахариды. В настоящее время с помощью электронного микроскопа установлено, что начальные стадии фибриллогенеза протекают в цитоплазме фибробластов, а заканчивается формирование коллагеновых волокон уже рядом с ними в окружающем основном межклеточном веществе.
Предполагаемый процесс образования фибробластами коллагеновых волокон представлен схематически на рисунке 48В. Синтезируемые на рибосомах молекулы белка тропоколлагена, пройдя через гранулярный эндоплазматический ретикулум и пластинчатый комплекс, выделяются в межклеточное основное вещество, где полимеризуются в крупные молекулы коллагена. Последние, укладываясь параллельно друг другу, объединяются в коллагеновую протофибриллу. В связи с усиленным биосинтезом тропоколлагена фибробласты обладают развитым гранулярным эндоплазматическим ретикулумом, а наличие в их цитоплазме мукополисахаридов связано с участием этих клеток в образовании основного межклеточного вещества. Фибробласты способны к движению скольжением и к сокращению, но оба эти процесса слабо выражены.
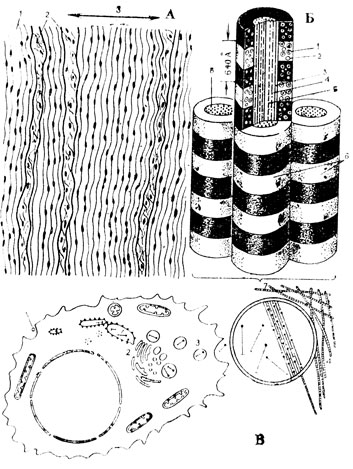
Рис. 48. А и Б - плотная волокнистая соединительная ткань коллагенового типа - сухожилие (продольный разрез). А - в оптическом микроскопе: 1 - пучок первого порядка; 2 - прослойки рыхлой соединительной ткани между пучками второго порядка; 3 - пучок второго порядка. Б - схема субмикроскопического строения коллагеновых волокон. Коллагеновое волокно (7) состоит из фибрилл (6). Осевая часть фибриллы (8) образована колластромином, в который входят белок аргирофильного типа (5), белок коллагенового типа (4) и полисахариды (5). Периферическая зона коллагеновой фибриллы образована проколлагеновым комплексом, состоящим из проколлагена (1) и полисахаридного компонента (2). В - схема образования коллагеновых фибрилл по Шелдону. В левой части рисунка молекула аминокислоты - пролина (1) захватывается фибробластом и включается в белок тропоколлаген. синтезируемый на рибосомах. Пройдя через гранулярную эндоплазматическую сеть и комплекс Гольджи (2), молекулы тропоколлагена (3) выделяются в межклеточное основное вещество, где они полимеризуются в более крупные молекулы коллагена. Последние соединяются в коллагеновую протофибриллу (это показано в двойном круге в правой части рисунка) с характерной исчерченностью (4)
Временно оседлыми клетками рыхлой волокнистой соединительной ткани являются гистиоциты. Их называли раньше "блуждающими клетками в покое", а теперь обозначают как "оседлые макрофаги". Каждое из наименований отражает определенную особенность данных клеток. Эти клетки идентичны подвижным или свободным макрофагам, только находятся в состоянии покоя.
Как мы уже указывали, свободные макрофаги могут трансформироваться и из разных других клеток - ретикулярных, лимфоцитов, моноцитов, а также из эндотелиальных клеток капиллярных расширений - синусоидов и выстилки синусов кроветворных органов, печени и некоторых других. Все вместе они образуют одну защитную функциональную систему, рассредоточенную по всему организму. Эту систему называют макрофагической (по И. И. Мечникову) или ретикуло-эндотелиальной, а сокращенно - РЭС. Все элементы этой системы способны к клеточным превращениям, фагоцитозу и нефрофагоцитозу.
Макрофаги, в отличие от фибробластов, обладают темными ядрами. Плазмолемма макрофагов характеризуется наличием складок и микроворсинок, облегчающих процесс фагоцитоза, а в цитоплазме, помимо всех клеточных органоидов, имеются многочисленные пиноцитозные пузырьки, лизосомы и фагосомы. Присутствие в цитоплазме макрофагов мукополисахаридов свидетельствует и об их участии в образовании основного межклеточного вещества соединительной ткани.
Макрофаги несут в организме еще одну важную функцию - участвуют в выработке антител.
Основной процесс выработки антител протекает в лимфоидной ткани, т. е. ткани органов лимфоидного кроветворения - лимфатических узлов и селезенки. При захвате антигенов (путем фагоцитоза) ретикулярными клетками, чаще макрофагами, информация об этом передается в органы системы иммуногенеза лимфоцитами, которые начинают дифференцироваться в плазматические клетки, или плазмоциты. Последние и являются продуцентами антител. Таким образом, функция антителообразования непосредственно принадлежит в организме высших позвоночных и человека плазмоцитам. Однако она сопряжена с подсобной ролью других клеток системы иммуногенеза и в первую очередь макрофагов.
Плазмоциты (рис. 13) морфологически отличаются от других клеток рыхлой соединительной ткани эксцентричным расположением ядра и наличием в цитоплазме светлого "дворика", примыкающего к ядру. В нем залегают пластинчатый комплекс и центриоли.
Цитоплазма этих клеток резко базофильна, так как содержит очень много рибосом. Эндоплазматический ретикулум плазмоцитов сплошь усеян рибосомами и может служить хорошим примером эргастоплазмы. Все это понятно в связи с интенсивным синтезом плазмоцитами антител -веществ белковой природы.
Помимо свободных макрофагов и плазмоцитов в рыхлой соединительной ткани встречаются еще другие подвижные клетки - тучные и лейкоциты крови, покинувшие кровяное русло.
Тучные клетки (лаброциты) содержат многочисленные зерна размером 0,3 - 1 мк, сходные по окрашиваемости с зернами базофилов, проявляющие, как и те, метахромазию. Резкая базофилия этих гранул обусловлена содержанием в них разных мукополисахаридов - гиалуроновой и хон-дроитинсерной кислот, а также гепарина. Ядро в тучной клетке относительно невелико, а органоиды слабо развиты. Клетками - предшественниками тучных клеток считают промакрофаги моноцитарного происхождения, сами же они редко делятся. Наличие в тучных клетках гепарина, который они могут выбрасывать из своего тела вместе с гранулами, позволяет думать, что они принимают участие в предотвращении свертывания крови. Однако до сих пор вопрос о функциональном значении тучных клеток не разрешен. По мнению исследователей, специально занимающихся изучением тучных клеток, секреторная функция этих элементов в свете данных последних лет представляется недостаточно обоснованной и вызывает серьезные сомнения. В гранулах тучных клеток были обнаружены гистамин и серотонин. Это находит свое объяснение в трактовке тучных клеток как особой разновидности макрофагальных элементов, специализированной на захвате, связывании и инактивировании указанных биогенных аминов (гистамина и серотонина). Тучные клетки являются высокодифференцированными зрелыми формами и, подобно гранулоцитам крови, ни к каким трансформациям не способны.
В рыхлой соединительной ткани могут встречаться еще округлые жировые и отростчатые пигментные клетки - хроматофоры (рис. 12), содержащие в цитоплазме гранулы бурого пигмента меланина. Жировые клетки (липоциты) обычно содержат одну жировую каплю, а цитоплазма имеет вид тонкого ободка вокруг нее. Небольшое утолщение этого ободка наблюдается в месте залегания ядра. Жировые клетки располагаются чаще по ходу кровеносных сосудов, по их сторонам. Разновидности рыхлой соединительной ткани, богатые скоплениями жировых клеток или густо заселенные хроматофорами, получили соответственно название жировой и пигментной ткани. Пигментная соединительная ткань образует одну из оболочек (пигментную) глаза и создает в его стенке темный экран, непроницаемый для световых лучей. Жировая соединительная ткань образует подкожную жировую клетчатку, жировые скопления в сальнике, около почки и в других местах тела. Жировая ткань играет в организме роль жирового депо, являющегося в случае недостатка пищи источником энергии, необходимой для поддержания жизни. За счет подобных жировых запасов протекают процессы жизнедеятельности во время зимнего сна у медведя или спячки у других животных. У морских зверей, в особенности у китообразных, мощно развитый жировой подкожный слой выполняет также важную функцию защиты тела от потери тепла, что позволяет этим организмам обитать в холодных водах. Механическое значение жировой ткани особенно наглядно выступает в жировой "сандалии", одевающей кости стопы. Благодаря множеству сконцентрированных здесь мелких "гидростатических" подушечек амортизируется давление большой тяжести нашего тела на точки опоры стопы. Без подобной жировой подстилки мы бы постоянно испытывали при стоянии на подошвах такую же боль, как и при долгом стоянии на коленях, где нет такого амортизатора. Интересно, что, не отличаясь по своему виду от обычных, жировые клетки жировой "сандалии" обладают специфической детерминированностью, т. е. определенной закрепленностью свойств, приобретенных в эмбриогенезе. Было установлено, что при предельном истощении человеческого организма, приводящем к смерти, когда в других местах тела все жировые резервы оказываются исчерпанными и жизнь поддерживается за счет других тканей, таких, например, как скелетная мускулатура, жировая "сандалия" стопы практически не изменяется, ибо это особая разновидность жировой ткани, несущая механическую функцию.
Была обнаружена (А. А. Браун) особая разновидность рыхлой соединительной ткани с отчетливо выраженной механической функцией. Это соединительнотканая основа кожи гренландской акулы. Данная ткань состоит из волокнистого каркаса - пучков коллагеновых волокон, расположенных пластами, параллельными поверхности кожи. Пласты залегают на большом расстоянии друг от друга, и эти промежутки в основном заполнены студневидной массой из мукополисахаридов, пронизанной тонкими волоконцами и тонкоотростчатыми фибробластами. Расположение волокнистых структур в этой ткани строго закономерное - в каждом пласте пучки лежат строго параллельно друг другу, а в смежных пластах ход их перекрещивается. Относительно тела рыбы все эти пучки идут диагонально, что обусловлено характером его движения. Эту ткань по морфологическому строению следует отнести к особой разновидности соединительной ткани - рыхловолокнистой оформленной.
Плотная волокнистая соединительная ткань выполняет в организме в основном механическую функцию и в большинстве своем является закономерно оформленной. Она встречается в двух видах - либо с преобладанием коллагеновых пучков (плотная соединительная ткань коллагенового типа, или фиброзная) или же с доминированием эластических волокон (плотная соединительная ткань эластического типа). Последняя имеет более однообразное строение. Она входит в состав желтых эластических связок, соединяющих кость с костью. Из этой ткани состоит, например, выйная связка, соединяющая затылочную кость черепа с шейными позвонками (рис. 47В). Благодаря растяжимости и эластичности волокнистых элементов этой связки, кости могут изменять свое положение относительно друг друга, а затем легко возвращаются в исходное состояние. И здесь эластические волокна представляют собой сплошные под световым микроскопом гомогенные тяжи, соединяющиеся друг с другом своими разветвлениями, и вся система благодаря этому имеет вид сети с удлиненными и узкими петлями. Большие диагонали указанных петель направлены по линиям натяжения. В промежутках между толстыми эластическими волокнами располагаются тончайшие прослойки рыхлой соединительной ткани, окружающие со всех сторон каждое эластическое волокно. По ходу же кровеносных сосудов, питающих эту ткань, прослойки рыхлой соединительной ткани приобретают значительные размеры. В рыхлой соединительной ткани, пронизывающей плотную эластическую, встречаются все клеточные формы, которые характерны для этой ткани.
Фиброзная ткань имеет несколько разновидностей. Наиболее просто построена плотная оформленная соединительная ткань коллагенового типа, из которой состоят сухожилия. Сухожилия - это органы, соединяющие мышцу с костью. Сократимой частью данной системы является мышечная ткань. Сухожилие же играет роль каната. Оно построено из прочных, но мало растяжимых коллагеновых пучков, ориентированных параллельно друг другу по линиям натяжения, то есть вдоль сухожилия (рис. 48А). Фибробласты и фиброциты, называемые здесь сухожильными клетками, залегают цепочками друг за другом, отделяя таким образом один от другого коллагеновые пучки первого порядка. Благодаря сдавливанию коллагеновыми пучками, сухожильные клетки приобретают своеобразную форму. Группы коллагеновых пучков первого порядка объединяются в пучки второго порядка, между которыми имеются хорошо выраженные прослойки рыхлой соединительной ткани, обозначаемые как эндотеноний. Пучки второго порядка подобным же способом объединяются в пучки третьего порядка и т. д., в зависимости от размеров органа. Общая оболочка, охватывающая сухожилие, носит название перитенония. В прослойках рыхлой соединительной ткани проходят сосуды и нервы, здесь же залегают и камбиальные элементы. Однако к делению способна и большая часть самих сухожильных клеток.
Более сложное строение имеют фасции, покрывающие мышцы. Они состоят из двух пластов коллагеновых пучков, плотно прилегающих друг к другу. В каждом из пластов ход пучков параллелен, в соседних он перекрещивается. По отношению к мускулу, его главной оси, волокна в обоих пластах идут наискось, что является приспособлением фасции к меняющейся толщине и длине мышцы. Часть коллагеновых пучков переходит из одного пласта в другой, скрепляя их таким образом. Здесь также имеются эластические волокна, образующие сети.
В организме встречается и неоформленная плотная соединительная ткань коллагенового типа. Многие исследователи относят к ней дерму кожи млекопитающих и человека. Однако для некоторых млекопитающих, в частности морских зверей, бегемота, свиньи, крупного рогатого скота, показана сложная закономерность хода коллагеновых пучков кожи и их переплетений. У других млекопитающих и у человека, обладающих большой гибкостью движений тела, связь коллагеновых пучков дермы приобретает настолько большую сложность, что ткань представляется нам неоформленной.
В пользу закономерного расположения волокнистых структур говорят так называемые лангеровские линии, обнаруживаемые в коже человека. Так, если нанести на человеческий труп в разных местах небольшие круглые дефекты во всю толщину кожи, то они сами по себе приобретают вид щелей и таким пунктиром обозначаются линии натяжения в каждом топографическом участке кожного покрова. Знание их важно хирургу для того, чтобы разрезы при оперативных вмешательствах производить в соответствии с этими линиями и тем способствовать более быстрому и полноценному заживлению ран.
Другой крайней группой тканей внутренней среды, противостоящей группе трофических тканей, являются развивающиеся из скелетогенной мезенхимы скелетные ткани, к которым относятся хрящ и кость.
Основную разновидность хрящевой ткани представляет стекловидный, или гиалиновый хрящ (рис. 49А). Это хрящ ребер и суставных поверхностей костей. Хрящевая ткань, как и костная, выполняет в организме прежде всего опорную функцию. В соответствии с ней построен, в частности, гиалиновый хрящ. Снаружи он одет надхрящницей, или перихондром, напоминающей в своей наружной зоне плотную соединительную ткань коллагенового типа. Клетки ее менее плотной, хондрогенной зоны представляют собой хондробласты, детерминированные как хрящевые и являющиеся камбиальными элементами данной ткани. Хрящевые клетки, расположенные в собственно хрящевой ткани, называют уже хондроцитами. Они не имеют отростков, чаще всего обладают округлой, треугольной или несколько вытянутой формой.
В отличие от хондробластов хондроциты окружены капсулами, обозначаемыми как хрящевые. По мнению сторонников образования основного вещества тканей внутренней среды путем эктоплазматического метаморфоза, хрящевая капсула - эктоплазма хрящевой клетки. Во всяком случае, этот слой основного вещества хряща, образующий стенку полости, в которой залегает хондроцит, обладает по сравнению с остальным межклеточным веществом иными физическими и химическими свойствами. Хондроциты возникают, во-первых, в результате размножения хондробластов и наложения молодых хрящевых клеток на предсуществовавшую ткань. Такой рост хряща называют аппозиционным. Во-вторых, происходит деление более молодых и менее дифференцированных хондроцитов в толще самого хряща. В результате образуются изогенные (то есть одного, общего происхождения) группы хрящевых клеток, содержащие все большее количество клеточных элементов. Это приводит к вставочному росту хряща. Хондроциты образуют вокруг себя межклеточное вещество и благодаря этому как аппозиционный, так и вставочный рост хряща становятся особенно хорошо заметными. Основу межклеточного вещества хряща составляет нежная сеточка переплетающихся друг с другом тонких коллагеновых пучков, которые называют хондриновыми волокнами. Вся их масса залита аморфной субстанцией - хондромукоидом, то есть хондромукопротеидом, представляющим собой мукополисахарид - хондроитинсерную кислоту, вещество очень большой вязкости.
Оно-то и придает хрящевой ткани ее основное физическое свойство - высоко выраженную способность оказывать сопротивление давлению.
Хрящевые клетки сами образуют и хондриновые волоконца, и хондроитинсерную кислоту. Хондроциты имеют типичное для клетки электронномикроскопическое строение (рис. 49Б). Синтез белка, хондрина, происходит на рибосомах гранулярного эндоплазматического ретикулума. Новообразованный хондрин перемещается в пластинчатый комплекс, где соединяется с новообразованным в пластинчатом комплексе мукополисахаридом и выделяется из клетки. В пластинчатом комплексе происходит также соединение мукополисахарида - хондроитинсерной кислоты и с неколлагеновым белком в хондромукопротеид. Вакуоли пластинчатого комплекса, содержащие хондрин и хондромукопротеид, сливаются с плазмолеммой, в результате чего оба эти вещества оказываются выведенными из хондроцита. Все эти процессы обеспечиваются энергией, доставляемой митохондриями.
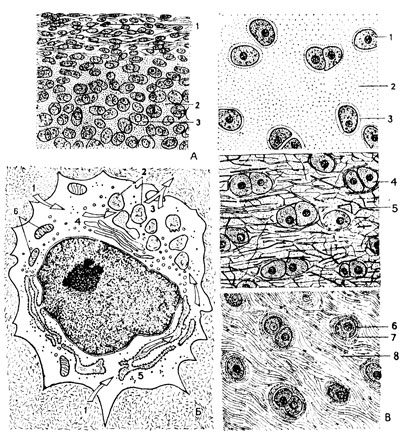
Рис. 49. Хрящевая ткань. А - молодой гиалиновый хрящ: 1 - надхрящница; 2 - межклеточное вещество; 3 - хрящевые клетки. Б - схема субмикроскопического строения хрящевой клетки (по Хэй): 1 - вещества, поступающие в клетку, из которых в ней строится межклеточная субстанция хряща. Синтез хондрина происходит на рибосомах гранулярного эндоплазм этического ретикулума (5). Новообразованный хондрин (коллаген хрящевой ткани) перемещается в пластинчатый комплекс (4), где соединяется с новообразованным в пластинчатом комплексе мукополисахаридом и выделяется из клетки (2 - 3). В комплексе Гольджи происходит также соединение мукополисахарида (хондроитинсерной кислоты) с неколлагеновым белком в хондромукопротеид. Вакуоли пластинчатого комплекса сливаются с плазмолеммой, в результате чего мукопротеиды и коллаген оказываются снаружи клетки. Все эти процессы обеспечивают энергией митохондрии (6) хрящевой клетки. В - разновидности хрящевой ткани: вверху - гиалиновая, в середине - эластическая, внизу - коллагенововолокнистая; 1 - хондроцит; 2 - межклеточное вещество; 3 - капсула хрящевой клетки; 4 - изогенная группа хондроцитов; 5 - эластиновая сеть; 6 - хрящевая клетка; 7 - вокругклеточное хрящевое основное вещество; 8 - пучки коллагеновых волокон
Участки межклеточного вещества, богатые хондроитинсерной кислотой, проявляют резкую базофилию и при окраске эозин-азуром интенсивно окрашиваются в синий цвет.
Молодое межклеточное вещество, прилежащее к хондроцитам, сравнительно бедно хондриновыми волокнами. Поэтому его базофилия выражена особенно сильно. Хрящевые же капсулы красятся слегка оксифильно, т. е. в данных условиях проявляют эозинофилию. Изогенные группы клеток, окруженные резко базофильными каемками, получили название клеточных полей, или клеточных территорий. В интертерриториальных пространствах значительно больше хондриновых, то есть коллагеновых структур, а коллаген, как известно, проявляет оксифилию. Поэтому участки между клеточными полями оказываются окрашенными в синий цвет менее интенсивно.
В старых хрящах у крупных животных либо в хрящах с резко выраженной механической функцией вокруг соседних изогенных групп образуется нечто вроде шарообразной коллагеновой капсулы, усиливающей прочность хряща. Такой ободок, естественно, проявляет в большей степени оксифилию, хоть и залит в какой-то мере кислым мукополисахаридом. Эти шаровидные образования являются как бы структурно-функциональными единицами подобной хрящевой ткани и получили наименование хондронов. В молодых хрящах не видно не только хондронов, но и разделение на клеточные поля и интертерриториальные пространства слабо выражено. Следует особо отметить, что хрящевая ткань, подобно пограничным тканям, лишена кровеносных сосудов и питание ее осуществляется путем диффузии тканевой жидкости через межклеточное вещество.
Кроме гиалинового хряща имеются еще 2 разновидности хрящевой ткани - эластическая и коллагенововолокнистая (рис. 49В).
Эластическая хрящевая ткань, образующая механическую основу ушной раковины, надгортанника, отличается от гиалиновой наличием пронизывающей ее межклеточное вещество сети эластических волокон. Она является опорой данного органа и в то же время придает ему большую эластичность.
Коллагенововолокнистая хрящевая ткань встречается главным образом между позвонками. Ее основа, построенная как гиалиновый хрящ, пронизана мощными коллагеновыми пучками, идущими параллельно друг другу (вдоль сил натяжения). Поэтому она напоминает по своему виду сухожилие, но вместо фиброцитов здесь находятся хондроциты и их изогенные группы, окруженные хрящевыми капсулами и более или менее значительными участками основного вещества. Подобные же структуры можно обнаружить на месте перехода сухожилия в кость, к которой оно прикрепляется опосредованно через волокнистый хрящ.
У старых животных и людей наблюдается обызвествление хряща - "асбестовая дистрофия" его межклеточного вещества.
Кость, как ткань, встречается в двух разновидностях - грубо- и тонковолокнистая, или иначе пластинчатая. Грубо-волокнистая кость присуща низшим позвоночным, а у высших встречается лишь в эмбриональном состоянии. Кость, как и хрящ, одета плотноволокнистой тканью - надкостницей, или периостом. В периосте, особенно в его глубоком и более тонковолокнистом слое, залегают многочисленные камбиальные клетки - остеобласты. Волокнистые структуры межклеточного вещества кости называются оссеиновыми, хотя по своей природе это коллагеновые волокна с присущей им оксифилией. Оссеиновые волокна впаяны в оссеомукоид - такой же мукополисахарид, как и хондромукоид. В отличие от хрящевой ткани, в костной волокна доминируют над мукополисахаридом. Главной же особенностью ее является богатство солями. В то время как хрящевая ткань содержит только около 2 - 3% солей (и их количество увеличивается лишь в старости), в кости неорганических веществ, например, столько же, сколько органических. Если взять именно костную ткань, а не кость как орган, то, по данным химического анализа, на соли приходится 28, а на оссеин - 30%. Соли эти теснейшим образом связаны с оссеином. Электронный микроскоп показывает, что мелкие кристаллики солей интимно спаяны с волокнистыми ее структурами. Соли кости - это различные соединения кальция. 85% приходится на фосфорнокислый кальций и 10% - на углекислый кальций.
О чрезвычайно интимной связи между оссеиновыми волокнами и солями кости можно судить по следующим фактам. Если, например, поместить кость на некоторое время в раствор кислоты, в результате чего из нее будут удалены соли, а затем изготовить гистологический препарат или такую же кость прокалить на огне и тем самым удалить из нее органические вещества, тонкая структура остатка кости в обоих случаях окажется одинаковой.
В грубоволокнистой кости оссеиновые пучки по своему расположению более всего напоминают переплет коллагеновых пучков в плотной неоформленной соединительной ткани коллагенового типа. Клетки, располагающиеся по ходу этих пучков, - костные клетки, или остеоциты, многоотростчаты (рис. 50Б). Их разветвляющиеся отростки пронизывают основное вещество. Сами тела клеток лежат в костных полостях, а отростки залегают в канальцах, пронизывающих межклеточное вещество. Стенки костных полостей и канальцев образуют в непосредственной близости к остеоцитам и их отросткам особые влагалища. Канальцы соседних клеток переходят друг в друга, а лежащие в них клеточные отростки анастомозируют. В щелевидных пространствах между остеоцитами и влагалищами циркулирует тканевая жидкость, обеспечивающая питание костным клеткам.
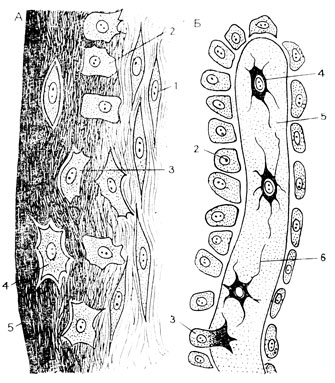
Рис. 50. Развитие кости из мезенхимы. А - образование межклеточного вещества (5) костной ткани остеобластами (2) и замуровывание последних (3) в это межклеточное вещество; 4 - костная клетка, лежащая в костной полости. Б - перекладина грубоволокнистой кости: 2 - остеобласт; 4 - костная клетка, остеоцит; 5 - межклеточное вещество; 6 - отросток остеоцита
Структурной единицей тонковолокнистой кости является пластинка (рис. 51), поэтому такую кость называют пластинчатой. Пластинка образована пучками оссеиновых фибрилл, склеенных оссеомукоидом. Пучки идут в пределах пластинки в одном направлении. В смежных пластинках ход их перекрещивается под разными углами, что придает костной ткани такого строения очень большую прочность. Остеоциты ориентированы вдоль хода пучков. Тела их располагаются как в толще пластинок, так и между ними. Подобное пластинчатое строение имеет как компактное, так и губчатое вещество костей скелета.
Наиболее сложно построено компактное вещество диафизов трубчатых костей. Основной структурной и функциональной единицей здесь являются остеоны, или гаверсовы системы пластинок.
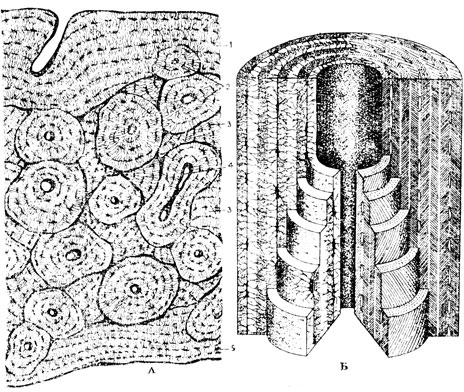
Рис. 51. Пластинчатая костная ткань. А - поперечный разрез через плотную (компактную) кость: 1 - слой наружных общих пластинок; 2 - остеоны; 3 - система вставочных пластинок; 4 - канал остеона; 5 - слой внутренних общих пластинок. Б - схема строения остеона
В остеоне пластинки имеют вид трубок, плотно насаженных друг на друга и ориентированных вокруг центрального канала; количество их колеблется от 4 до 20. По каналу остеона проходят тонкостенные кровеносные сосуды и нервы. Из кровеносных сосудов в костные канальцы и полости, пронизывающие пластинки, просачивается кровяная плазма, которая далее циркулирует по ним в виде тканевой лимфы, питающей остеоциты. Система эта замкнутая - канальцы наружной пластинки имеют аркоподобный вид и за пределы остеона не заходят. Сам остеон отграничен спаивающей полоской. Описанное строение остеонов обусловливает их большую прочность и способность противостоять давлению.
В трубчатой кости между остеонами залегают вставочные пластинки, а по периферии компактной кости диафиза снаружи и изнутри лежат общие, или генеральные пластинки (рис. 51).
На границе с костномозговой полостью к внутренним генеральным пластинкам прилежит эндост, представленный нежной соединительнотканной прослойкой. Как и периост, эндост богат остеобластами.
Кровеносные сосуды, васкуляризирующие компактное костное вещество, проникают в него со стороны надкостницы через питательные каналы, пронизывающие наружные генеральные пластинки. Надкостница спаяна с компактным костным веществом с помощью прободающих коллагеновых волокон, вмурованных на разную глубину в наружные генеральные пластинки.
Как выяснилось из изучения структуры костей в динамике, костная ткань все время перестраивается в соответствии с изменяющимися механическими условиями нагрузки на кость как орган. В этом процессе перестройки одни остеоны разрушаются, а другие строятся заново. Вставочные пластинки - это остатки бывших остеонов. На месте любого из имеющихся в данный момент остеонов со временем может оказаться лишь след в виде участка пластинок, которые будут как бы вставлены между новообразованными остеонами. То же относится и к губчатому костному веществу, из которого, например, почти нацело (за исключением тонкого наружного слоя компактной кости) построены эпифизы трубчатых костей. Его перекладины образованы пластинками тонковолокнистой костной ткани (рис. 52-12 и 14). В их расположении наглядно выступают органные закономерности, ибо оно всегда отвечает наиболее полноценному выполнению механической функции (противостоянию давлению и растяжению) при наименьшей затрате материала.
Мы рассмотрели строение грубоволокнистой и пластинчатой кости как ткани, а затем строение трубчатой кости как органа. Разберем теперь вопрос о развитии кости.
Развитие грубоволокнистой костной ткани из мезенхимы наблюдается очень рано, уже в первый месяц эмбриогенеза. Процесс начинается с активной пролиферации мезенхимных клеток и появления остеогенных островков. Затем в этих остеогеиных островках возникают очажки новообразования коллагеновых волоконец, которые позднее спаиваются вместе оссеомукоидом. И то, и другое продуцируют мезенхимные клетки, дифференцирующиеся в остеобласты и частично замуровывающиеся в разрастающуюся костную ткань (рис. 50А). В результате образуются перекладины грубоволокнистой кости с остеоцитами, заточенными в костные полости, а вокруг этих балочек остеобласты сидят настолько густо, что слой их приобретает эпителиообразный вид. Остеобласты - крупные клетки, эазмером до 20 мк, ядра у многих лежат эксцентрично. Как у всех клеток, интенсивно синтезирующих белок, у остеобластов, образующих межклеточное вещество кости, сильно развит гранулярный эндоплазматический ретикулум. Хорошо выражены комплекс Гольджи и митохондрии. Процесс новообразования оссеиновых волокон, скрепления их оссеомукоидом и вмуровывания в оплотневающее межклеточное костное вещество остеобластов (с дальнейшим превращением их в высокодифференцированные остеоциты, уже не способные к делениям) продолжается. Наконец, происходит обызвествление межклеточного вещества и грубоволокнистая костная ткань оказывается полностью сформированной.
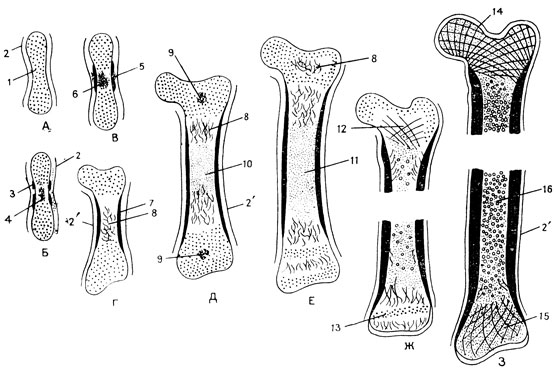
Рис. 52. Развитие кости на месте хряща. А - кость как орган представлена хрящевой моделью (1), 2 - надхрящница. Б - вокруг хрящевой модели образовалась перихондральная костная манжетка (3), а в середине диафиза хрящевой модели появилась первичная точка обызвествления, или окостенения (4). В - через перихондральную костную манжетку в точку окостенения прорвалась остеогенная мезенхима (5), разрушающая обызвествленный хрящ и на остатках его строящая эндохондральную кость (6). Г - перихондральная (7) и эндохондральная (5) костная ткань разрастается в стороны эпифизов, 2' - надкостница. Д - эндохондральная кость в средней части диафиза разрушается и начинается образование костномозговой полости (10), в эпифизах появляются вторичные точки обызвествления, или окостенения (9), 2' - надкостница. Е - начинается образование костного мозга. Ж - появляются первые пластинки губчатого костного вещества в эпифизе (12); возникает хрящевая пластинка роста (13) между эндохондральной костной тканью диафиза и эпифиза. 3 - перекладины имеют строго закономерное расположение (14), в губчатом веществе эпифизов располагается миелоидная кроветворная ткань - красный костный мозг (15), в диафизе залегает желтый костный мозг, состоящий главным образом из жировой ткани (16)
Развитие трубчатой кости как органа происходит на месте хряща, точнее на месте ее хрящевой модели, образовавшейся из мезенхимы (рис. 52А). Оно начинается на втором месяце эмбриогенеза с образованием в центре диафиза костной манжетки (перихондрального костного кольца). Она образуется остеобластами надхрящницы и состоит из перекладин грубоволокнистой кости, которые связаны друг с другом в сеть. Образование перихондральной манжетки нарушает питание хрящевой модели в этой части диафиза. Это приводит к дистрофическим изменениям хряща и его обызвествлению в данном месте. Так появляется в центральной части диафиза хрящевой модели первичная точка обызвествления, или окостенения.
Через промежутки между перекладинами перихондральной костной манжетки в точку обызвествления (окостенения) хрящевой модели проникает остеогенная мезенхима и, разрушая обызвествленный хрящ, строит на его остатках костную ткань. Так образуется внутрихрящевая, или эндохондральная кость.
Перихондральная кость разрастается под надкостницей вокруг хрящевой модели. Процесс обызвествления хряща продвигается по направлению к эпифизам, вслед за этим происходит разрушение новых участков обызвествленного хряща и замещение его эндохондральной костью. Одновременно возникают в эпифизах вторичные точки обызвествления, или окостенения хряща, а в центральной части диафиза наблюдается разрушение эндохондральной кости и начало образования костномозговой полости. Разрушение костной ткани осуществляется особыми клетками "костедробителями" - остеокластами. Это многоядерные клетки крупных размеров (до 100 мк). В их цитоплазме имеется много митохондриев и лизосом, богатых гидролитическими ферментами. С помощью этих ферментов остеокласты разрушают межклеточное вещество кости.
Далее происходит эндохондральное окостенение в эпифизах. Костномозговая полость в диафизе удлиняется и в ней проникшая сюда мезенхима строит костный мозг. В результате разрастания навстречу друг другу эндохондриальной кости из эпифиза и диафиза область, занятая хрящевой тканью, сокращается и остается только хрящевая пластинка роста, за счет которой продолжается удлинение кости. В эпифизах начинается образование пластинок губчатой кости. Перихондральная, точнее, уже периостальная кость сильно разрастается в длину и увеличивается в толщину. В ней происходит перестройка - грубоволокнистая костная ткань замещается пластинчатой. Грубоволокнистая кость разрушается остеокластами, а пластинчатая костная ткань строится вокруг врастающих сюда кровеносных сосудов, ориентированных вдоль кости.
В эпифизах нарастает количество пластинок губчатой кости, а промежутки между ними оказываются заполненными миелоидной тканью. Костномозговая полость в области диафизов содержит уже желтый костный мозг, то есть жировую ткань, развившуюся за счет ретикулярной ткани костного мозга. Компактная кость перихондрального происхождения приобрела уже свое дефинитивное строение.
Таким образом, в большую группу опорно-трофических тканей, объединенную общностью происхождения из мезенхимы, входят представители с различным функциональным значением и соответственно с разными структурными особенностями. На полюсах этой большой тканевой системы находятся такие противоположные по своему характеру ткани, как кровь и лимфа, с одной стороны, хрящ и кость - с другой. Однако и во взрослом организме отчетливо выражено единство всей системы тканей внутренней среды. Оседлая недифференцированная, адвентициальная клетка может при нормальном развитии, а тем более в эксперименте или в условиях патологии, дать начало всем видам оседлых клеточных элементов соединительной, хрящевой и костной тканей (фибробласту, жировой клетке, хондроциту, остеоциту). Из нее может начаться процесс кроветворения в любом участке соединительной ткани. Эта же клетка может дать начало макрофагам и тучным клеткам, а макрофаги в свою очередь могут трансформироваться в фибробласты.
Примитивная ретикулярная клетка может дифференцироваться во все те же клетки, что и адвентициальная. Кроме того, трансформировавшись в гемоцитобласт, она может послужить источником образования форменных элементов крови и плазматических клеток. Лимфоциты же могут в итоге превратиться в фибробласты. И наконец, следует вспомнить, что сам процесс кроветворения во взрослом организме вынесен в соединительную ретикулярную ткань, а между кровью и соединительной тканью происходит постоянный обмен клеточными элементами.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'