Глава VIII. Определение, классификация тканей и развитие их из эмбриональных зачатков
Самое краткое определение понятия ткань мы дали в начале книги. Там мы отметили, что ткани - это строительные материалы, из которых состоят органы. В свою очередь, ткани построены из клеток и их производных, но это не какие-то беспорядочные конгломераты, комплексы структурных инградиентов, а строгие системы, выполняющие в организме определенные элементарные функции - отграничения и избирательно проницаемого барьера, внутреннего обмена и опоры, сокращения и движения, восприятия и передачи раздражения. Сконструированные в соответствии с выполняемой ими работой ткани являются функциональными системами, в которых составляющие их клетки и клеточные производные взаимосвязаны друг с другом. Таким образом, более широким определением ткани будет понимание ее как функциональной системы клеток и их производных.
Однако и такое определение является неполным. В нем отсутствует момент развития. Между тем в определении ткани необходимо подчеркнуть, что все ее составные элементы связаны не только структурной и функциональной общностью, но и общим происхождением в онтогенезе, т. е. в индивидуальном развитии. Все клеточные элементы одной ткани развиваются из единого эмбрионального зачатка. С другой стороны, эволюционная гистология, т. е. такое направление в изучении тканей, которое разрабатывает вопросы закономерностей и путей их исторического развития, говорит о том, что ткани не оставались неизменными, а изменялись вместе с организмами в процессе филогенеза (развития вида). Особенность эволюционного развитии тканей состоит в том, что сами они являются лишь частями целого организма и потому непосредственно не подлежат действию естественного отбора - важнейшей движущей силы филогенетического развития живых существ. Ткани подвергаются отбору в процессе эволюции опосредованно через организм. Меняются условия существования, и в новых условиях среды выживают особи, наиболее приспособленные к ним, функции органов и тканей которых наилучшим образом отвечают требованиям, предъявляемым в новых условиях их существования. Отбор происходит по организмам, но в основе его лежат тканевые и органные изменения. Неудачные изменения элиминируются (отбрасываются), а удачные приводят в итоге к существенным преобразованиям тканей, переопределяющим их коренные особенности.
Учитывая все сказанное, мы можем предложить теперь полное определение ткани, в основу которого положены понимание их и формулировка, данная в свое время основоположником советской эволюционной гистологии академиком А. А. Заварзиным. Ткани - это исторически сложившиеся при взаимодействии организма с внешней средой функциональные системы клеток и их производных, объединенные общим происхождением в онтогенезе и детерминацией, т. е. потенциями развития.
Разбирая вопрос о дифференцировке клеток в процессе онтогенеза, мы отмечали, что, по определению В. П. Михайлова, ткань - "частная система организма, состоящая из клеток с одинаковым образом реализуемой эпигеномной наследственностью". Будучи по существу правильным, данное определение ткани является, однако, далеко не полным, так как не подчеркивает функционального значения тканевых структур, не указывает на их филогенетическую обусловленность и опускает наличие в ткани неклеточных компонентов, в то время как в нашем теле на надклеточные и межклеточные производные клеток приходится примерно 4/5 его массы. Введением в определение ткани их детерминированности, то есть закрепленности потенций развития, мы фактически охватываем и представление об одинаковым образом реализуемой ими их эпигеномной наследственности.
В соответствии с элементарными функциями (по А. А. Заварзину - константами), которые ткани выполняют в теле животных и человека - обеспечение пограничности, постоянства внутренней среды, движения, реактивности - их разделяют на 4 основные группы: 1) эпителиальные ткани с ведущей отграничивающей ролью; 2) ткани внутренней среды; 3) мышечные ткани, осуществляющие сократительную работу; 4) нервные ткани, клетки которых воспринимают раздражения, поступающие из внешней или внутренней среды, перерабатывают их в специфические импульсы и передают на те или иные рабочие структуры в виде соответствующих распоряжений. Основы этой классификации были заложены Келикером и Лейдигом более ста лет тому назад.
В настоящее время все многообразие конкретных тканей разных животных, за очень редким исключением, удается уложить в указанные четыре тканевые группы этой классификации. Особенно отчетливый морфофункциональный (т. е. основанный на подчеркивании функционального значения структуры, их единства) характер придал этой классификации академик А. А. Заварзин. Он предложил заменить термин "эпителиальные" на "пограничные", благодаря чему подчеркивается их значение как структур, лежащих на границе внутренней и наружной среды организма. Точно так же, употребляя вместо широко принятого названия "ткани промежуточного вещества" новое "ткани внутренней среды", А. А. Заварзин выдвинул на первый план ведущую функцию данных тканей, хотя, пожалуй, более точно характеризует роль большой и наиболее разнообразной группы этих тканей наименование "опорно-трофические".
Подробное описание каждой из перечисленных групп тканей будет дано в последующих главах книги. Здесь же мы считаем необходимым еще раз акцентировать внимание на ярко выступающей при изучении тканей диалектической закономерности единства структуры и функции, строения и работы данных систем. Она отчетливо проявляется, как мы увидим дальше, и в особенностях, отличающих друг от друга разновидности тканей в пределах каждой из этих четырех тканевых групп. Теперь же перейдем к вопросу о развитии тканей в онтогенезе.
Ткани, сложившиеся в процессе филогенеза организмов и наследственно закрепленные как таковые, формируются каждый раз заново на определенном этапе индивидуального развития из эмбриональных зачатков.
Эмбриональный период индивидуального развития, т. е. развитие зародыша, начинающееся после оплодотворения, можно условно разбить на следующие этапы: зигота (этап одноклеточного зародыша), дробление, гаструляция и органогенез.
Яйцевая клетка в процессе оплодотворения пассивна. Единственным незначительным проявлением ее активности является образование воспринимающего бугорка навстречу первому сперматозоиду, разрушающему все преграды на пути к ней. Сперматозоид, наоборот, весьма активен. Его движение к яйцу определяется двумя раздражающими факторами - током жидкости (устремляется навстречу) и, главное, хемотаксическим воздействием со стороны яйцеклетки (идет по направлению химических сигналов - продуцируемых ею метаболитов).
При оплодотворении в яйцеклетку внедряется не весь сперматозоид, а только головка, шейка и промежуточный отдел, хвостик отбрасывается. Яйцевая клетка, покинувшая яичник и проделывающая деления созревания уже в яйцеводе, способна к оплодотворению всего в течение 2 часов. Оплодотворение у высших позвоночных и человека внутреннее и происходит в верхней трети яйцеводов. Попав в воронку яйцеводов, зрелая яйцеклетка оказывается одетой слоем обслуживавших ее в процессе развития трофических фолликулярных клеток. Спаянные друг с другом мукополисахаридом - гиалуроновой кислотой, они образуют вокруг яйцеклетки лучистый венец (рис. 36), через который и должны пробиться сперматозоиды. Акросома, или перфораторий, выделяет фермент гиалуронидазу, который разрушает гиалуроновую кислоту, цементирующую слой фолликулярных клеток. Только совокупным действием множества сперматозоидов может быть преодолен этот барьер. Вот почему для осуществления акта оплодотворения необходима определенная концентрация в сперме живчиков.
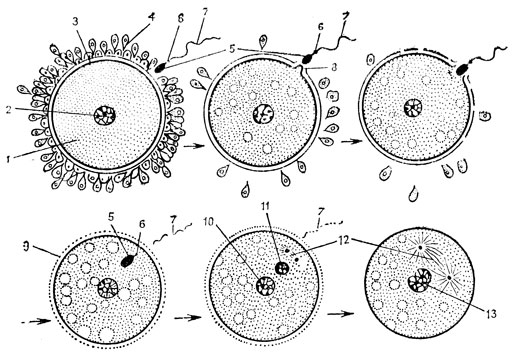
Рис. 36. Оплодотворение (из А. Г. Кнорре): 1 - яйцо; 2 - его ядро; 3 - блестящая оболочка; 4 - лучистый венец из фолликулярных клеток; 5 - головка сперматозоида; 6 - его шейка; 7 - хвостик; 8 - воспринимающий бугорок; 9 - желточная оболочка; 10 - женский пронуклеус; 11 - мужской пронуклеус; 12 - ахроматиновое веретено между центриолями; 13 - формирующийся синкарион
В эякуляте, попадающем при семяизвержении в половые пути женщины, имеются многие миллионы сперматозоидов. Однако в яйцеклетку входит только один сперматозоид. Как только один из живчиков, атакующих женскую половую клетку, попадает в нее, на поверхности яйцеклетки образуется желточная оболочка-мембрана оплодотворения, препятствующая проникновению в нее других сперматозоидов. Сперматозоид вносит в яйцеклетку все свои органоиды, в том числе и клеточный центр. Проксимальная и дистальная центриоли начинают расходиться к полюсам зиготы, и это является уже подготовкой к делению оплодотворенной яйцеклетки на две дочерние клетки - два бластомера (по-гречески "бласте" - зародыш, а "мерос" - часть). Головка сперматозоида, т. е. его ядро, разбухает, увеличивается в размерах и продвигается к центру зиготы, где лежит материнский пронуклеус. Оба пронуклеуса сливаются в одно соядрие - синкарион. К этому времени центриоли оказываются на полюсах зиготы и между ними образуется ахроматиновый митотический аппарат. В синкарионе из его пронуклеусов оформляются хромосомы - в каждом их гаплоидное количество, а вместе - диплоидное. В хромосомах становятся хорошо заметными хроматиды и сразу, таким образом, формируется экваториальная пластинка метафазы. Далее сестринские хроматиды расходятся по дочерним бластомерам и каждый из них получает одинаковый диплоидный набор хромосом, т. е. полный набор как материнских, так и отцовских генов.
На следующем этапе - дроблении - митотические деления бластомеров быстро следуют друг за другом. В интерфазе не происходит увеличения размеров дочерних клеток до размеров материнской, и материал зиготы все более и более раздробляется на уменьшающиеся по величине бластомеры. В зависимости от степени загруженности зиготы желтком дробление может быть полным или неполным, частичным, например, у птиц, где яйцеклетки переполнены желточными зернами и дроблению подвергается лишь незначительная часть зиготы, имеющая вид диска и получившая поэтому наименование зародышевого диска. Полное дробление встречается в двух вариантах. Если в зиготе совсем мало желтка и он не мешает процессу дробления (это имеет место у ланцетника), последнее носит равномерный характер и бластомеры соизмеримы друг с другом по величине (рис. 37). Если в зиготе много желтка, например, у земноводных, дробление оказывается полным, но неравномерным - бластомеры нижнего, вегетативного полушария значительно крупнее верхних (рис. 38), анимальной половины зародыша. Последние бластомеры делятся активнее и количество клеток на анимальном полушарии оказывается в итоге большим, чем на вегетативном.
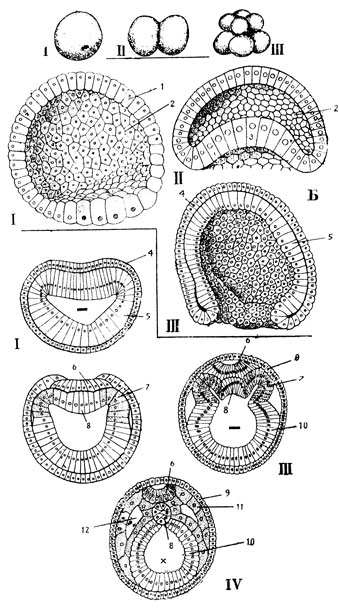
Рис. 37. Эмбриональное развитие ланцетника. А - период дробления: I - зигота, II - двухбластомерная стадия, III - восьмибластомерная стадия. Б - бластула (I); ранняя (II) и более поздняя (III) стадии гаструляции: 1 - бластодерма; 2 - бластоцель (первичная полость тела); 3 - начало впячивания (инвагинации) вегетативной части бластулы; 4 - первичная эктодерма; 5 - первичная энтодерма. В - закладка осевых органов. Поперечные разрезы ранней (I) и более поздней (II) гаструл, ранней (III) и поздней (IV) нейрул: 6 - нервная пластинка, прогибающаяся затем в нервный желобок; 7 - мезодермальные пузыри; 5 - хорда; 9 - кожная эктодерма; 10 - кишечная энтодерма; 11 - первичный сегмент мезодермы; 12 - вторичная полость тела (целомическая). Знаком '-' показана полость первичной кишки (гастроцель), знаком 'x' - полость кишечной трубки, в которую замкнулась кишечная энтодерма
Первая борозда дробления у всех вышеназванных животных проходит меридионально, вторая борозда тоже меридиональная, но ложится перпендикулярно первой. При полном и равномерном дроблении третья борозда почти экваториальная, при полном неравномерном она широтная и располагается значительно выше экватора. Затем чередуются друг с другом меридиональные и широтные борозды, причем деления клеток протекают синхронно, одновременно, как по команде. У земноводных и птиц к меридиональным и широтным бороздам дробления затем добавляются еще и тангенциальные борозды (т. е. параллельные тангенсу, касательной к поверхности зародыша).
В итоге этапа дробления возникает бластула. У ланцетника бластула представляет собой пузырь со стенкой из одного слоя клеток, называемой бластодермой (рис. 37Б). Полость пузыря носит название первичной полости тела, или бластоцеля. У амфибий бластула тоже пузыревидная, но бластодерма ее образована несколькими слоями клеток (рис. 38-6 и 39-1). У птиц бластула имеет дисковидную форму и залегает на желтке, представляющем собой массу питательного материала для зародыша. У млекопитающих дробление полное, поскольку в яйцеклетке очень мало питательных веществ и они необходимы лишь на самое первое время зародышевого развития, а эмбриогенез протекает в основном в особом органе - матке, обеспечивающем зародыш всем необходимым для его развития. Однако дробление у них происходит асинхронно. В итоге этапа дробления у млекопитающих, как и у птиц, образуется дискобластула.
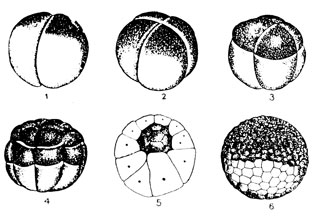
Рис. 38. Эмбриональное развитие амфибий: 1-5 - дробление; 6 - бластула
Как показали опыты с маркировкой, т. е. нанесением цветных меток прижизненными красителями на разные участки бластулы, на этой стадии эмбриогенеза уже можно сказать, какая область ее даст при нормальном развитии начало той или иной тканевой закладке. Фогт, например, установил на амфибиях, что в анимальном полушарии бластулы размещаются материалы будущего кожного эпителия - кожной эктодермы и рядом с ним будущей нервной ткани - нервной пластинки, а на вегетативном полушарии залегает материал будущего кишечного эпителия - кишечной энтодермы. Между ними располагается пояс из хорды и, главным образом, мезодермы с зачатками других тканей (рис. 40-1). Однако все эти закладки являются презумптивными, т. е. предположительными. Они еще не закреплены, не детерминированы как таковые и это легко показать экспериментами с перемещением кусочков стенки бластулы с места на место. Если, например, поменять местами небольшие участки из области будущей кожной эктодермы и будущей нервной пластинки и проследить за развитием зародыша, никаких уродств мы не обнаружим.
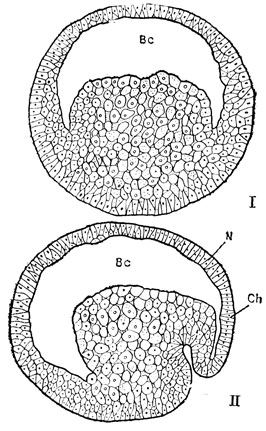
Рис. 39. I - разрез бластулы амфибий, II - начало гаструляции, инвагинации. BC - бластоцель, полость бластулы (первичная полость тела); N -материал нервной пластинки; Ch - материал хорды
Следующий этап эмбрионального развития - гаструляция состоит в образовании зародышевых листков, в размещении по этим листкам указанных эмбриональных закладок и в их расположения детерминации. У ланцетника зародыш из однопластового становится двухслойным, а затем трехслойным (рис. 37В) У амфибий из бластулы в процессе гаструляции (рис. 39, 40, 41 и 42) сразу образуется зародыш. Мы ограничимся рассмотрением гаструляции лишь у ланцетника и земноводных, протекающей более просто, чем у высших позвоночных, где этот процесс осложняется образованием зародышевых оболочек, изолирующих эмбрион от внешней среды и обеспечивающих его необходимыми питательными веществами.
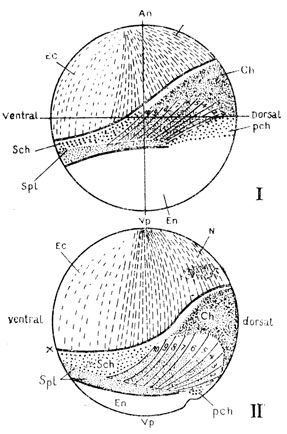
Рис. 40. Карта расположения презумптивных закладок у амфибий в бластуле (I) и в начале гаструляции (II). An - анимальный полюс; Vp - вегетативный полюс; Ec - кожная эктодерма; N - нервная пластинка; Ch - хорда; En - кишечная энтодерма; pch - часть ее, обозначаемая как прехордальная пластинка (при развитии дает начало головной кишке); части мезодермы: 1 - 10 - сомиты, Spl - закладка боковых пластинок (спланхнотома), Sch - закладка хвоста, x - x - граница подворачивания мезодермы и хорды
В классической работе А. О. Ковалевского (1861) по развитию ланцетника было показано, что путем впячивания, инвагинации стенки бластулы зародыш на стадии ранней гаструлы оказывается состоящим из наружного и внутреннего листков. Эти эпителиобразно построенные пласты клеток получили соответственно наименование первичной эктодермы и первичной энтодермы. Впяченным оказывается вегетативное полушарие бластулы. Полость гаструлы обозначают как гастроцель, первичную кишку, а отверстие, ведущее в гастроцель, как бластопор, или первичный рот. Остаток бластоцеля (первичной полости тела) представлен щелью между первичной эктодермой и первичной энтодермой.
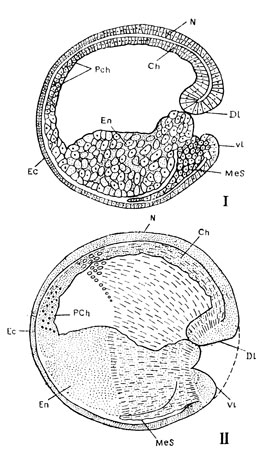
Рис. 41. Медиальный разрез гаструлы амфибии. На рисунке II область распространения мезодермы (Mes) между наружным и внутренним зародышевыми листками нанесена штрихами. Dl - дорзальная, Vl - вентральная губы бластопора. Остальные обозначения те же, что и на предыдущем рисунке
Согласно описаниям А. О. Ковалевского, внутренний пласт - первичная энтодерма образует симметрично в спинной, дорзальной своей части выпячивания - пузыри, дающие начало среднему зародышевому листу, или мезодерме, и зародыш становится трехслойным, трехлистковым. Пузыри эти потому и получили название мезодермальных. Располагаются они метамерно, как бы члениками вдоль тела зародыша. Участок первичной энтодермы между мезодермальными пузырями превращается в хорду - осевую струну эмбриона. Оставшаяся брюшная - вентральная часть первичной энтодермы смыкается в трубку, которая теперь уже является кишечной энтодермой. В первичной эктодерме над хордой клетки приобретают большую высоту и в совокупности образуют нервную пластинку. Позднее, уже на последнем этапе эмбриогенеза - органогенезе нервная пластинка прогибается в нервный желобок, отрывается от эктодермы и заворачивается в нервную трубку. После выделения нервной пластинки первичная эктодерма становится кожной эктодермой. Впоследствии кожная эктодерма смыкается над нервной пластинкой, прогибающейся в нервный желобок. На этой стадии зародыш ланцетника оказывается состоящим из пяти эмбриональных закладок, собранных в три зародышевых листка. Осевое расположение хорды относительно тела зародыша, залегание по сторонам от нее мезодермы, а над ней нервной пластинки является типичным для всех хордовых животных.
Более поздние и детальные исследования процесса гаструляции у ланцетника подтвердили правильность описаний, сделанных А. О. Ковалевским.
У амфибий гаструляция происходит одновременно путем медленной инвагинации клеток вегетативного полушария и одновременного наползания, эпиболии мелких клеток анимального полушария. Место инвагинации - бластопор становится впоследствии анальным отверстием, а рот прорывается на противоположном конце тела (почему хордовые получили вместе с представителями некоторых других типов многоклеточных животных, в частности с иглокожими, наименование вторичноротых - Deuterostomia). Одновременно с инвагинацией материала кишечной энтодермы внутрь гаструлы у амфибий в области бластопора происходит вклинивание материала мезодермы между наружным и внутренним листками (рис. 41). В итоге гаструляции у амфибий возникает эмбрион, состоящий из трех зародышевых листков с таким же распределением по листкам эмбриональных закладок (рис. 42).
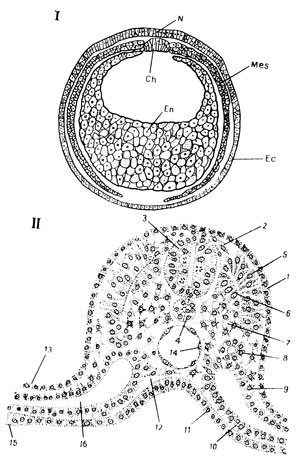
Рис. 42. 1 - поперечный разрез поздней гаструлы амфибий: N - нервная пластинка; Ch - хорда; Mes - мезодерма; Ec - кожная эктодерма, En - кишечная энтодерма. II - схема (А. Г. Кнорре) органогенеза у зародыша высшего позвоночного (названия тканевых производных поставлены в скобках после названия соответствующего зачатка: 1 - кожная эктодерма (эпидермис); 2 - ганглиозная пластинка (чувствительные нейроны, периферическая нейроглия, хроматофоры); 3 - нервная трубка (нейроны, нейроглия); 4 - хорда; 5 - дерматом (соединительнотканая основа кожи); 6 - миотом (скелетномышечная ткань); 7 - склеротом (хрящевая и костная ткани); 8 - нефротом (почечный эпителий); 9 - париетальный листок спланхнотома (мезотелий, ткань сердечной мышцы); 11 - кишечная энтодерма (кишечный эпителий; 12 - мезенхима (соединительная ткань, кровь, гладкомышечная ткань и, по мнению большинства исследователей, эндотелий); 13 - внезародышевая эктодерма (эпителий амниона); 14 - эндотелий аорты; 15 - желточная энтодерма (эпителий желточного мешка); 16 - целом (вторичная полость тела, впоследствии - брюшная, грудная и околосердечная полости)
На стадии поздней гаструлы материалы эмбриональных закладок уже оказываются детерминированными. Если теперь попробовать поменять местами участки материалов, например, из кожной эктодермы и нервной пластинки, то при дальнейшем развитии зародыша можно будет обнаружить соответствующие дефекты - в области кожи разовьется участок нервной ткани, а в пределах нервной системы - очаг кожного эпителия. Процесс детерминации эмбриональных закладок является результатом взаимного влияния их друг на друга. В дальнейшем весь ход эмбриогенеза определяется подобными коррелятивными взаимодействиями между развивающимися частями тела зародыша.
Наибольшие изменения на последнем этапе эмбрионального развития - органогенезе наблюдаются в среднем зародышевом листке (рис. 42). Прежде всего в вентральной части мезодермы появляется щель - вторичная полость тела, или целом, отграниченная снаружи пристеночным (париетальным), а изнутри внутренностным (висцеральным) листками. Вместе они представляют собой брюшной мешок, или спланхнотом. Дорзальная мезодерма имеет метамерный, членистый характер и состоит из спинных сегментов, или сомитов. Каждый сомит соединен со спланхнотомом сегментной ножкой, или нефротомом. Позднее спинные сегменты дифференцируются на три части: латеральную - дерматом, медиальную - склеротом и срединную - миотом. Еще позднее дерматом и склеротом начинают рассыпаться на отдельные клетки, соответственно на дермальную, кожную и скелетогенную мезенхиму. Из первой развивается соединительнотканая основа кожи, из второй - хрящевая и костная ткани скелета.
Мезенхимным клеточным элементам дают начало и другие участки мезодермы. Например, из висцерального и париетального листков спланхнотома начинают выселяться клетки, из которых формируется неспецифическая пристеночная и внутренностная мезенхима. Постепенно мезенхимные клетки заполняют все свободные промежутки между эмбриональными закладками, т. е. все остатки первичной полости тела. Свое наименование мезенхима первичная эмбриональная промежуточная ткань) получила от греческих слов "энхимо" - заполняю и "мезо" - середина.
Нефротомы обособляются сначала от спинных сегментов, затем от спланхнотомов и формируют мочевые канальцы. Миотомы разрастаются в дорзальном и вентральном направлениях и образуют скелетную мышечную ткань. Листки опланхнотома превращаются в эпителиальную выстилку вторичной полости тела - мезотелий. Кожная эктодерма развивается в эпидермис и его производные, кишечная энтодерма - в эпителиальную выстилку желудочно-кишечного тракта, а также в эпителий поджелудочной железы и печени. Висцеральная мезенхима строит как соединительнотканую основу, так и гладкомышечные элементы внутренних органов. Нервная пластинка изгибается в нервный желобок, который затем отрывается от кожной эктодермы и заворачивается в нервную трубку. Пограничные участки нервного желобка обособляются в парные образования, лежащие метамерно по сторонам нервной трубки - спинальные ганглии. В области туловища нервная трубка является закладкой спинного мозга, в головном конце зародыша она расширяется, образуя сначала 3, а потом 5 мозговых пузырей, которые дают начало разным отделам головного мозга. Из нейронов, расположенных в нервной трубке и спинальных ганглиях, вырастают дендриты и нейриты, которые разрастаются по всему эмбриону и устанавливают сложнейшие связи между самими нервными клетками, а также между нервными клетками и всеми частями тела зародыша.
Таким образом, мы видим, что по ходу развития зародыша имеет место все более детальная дифференцировка его эмбриональных зачатков. Развиваясь дивергентно, эмбриональные закладки в конце концов дают начало уже специфически функционирующим строительным материалам органов - тканям. Возникшие из эмбриональных закладок ткани приобретают стойкие качественные различия. Дифференцируясь, эмбриональные зачатки все более теряют способность взаимного превращения. Развившиеся из них и приобретшие прочную детерминированность ткани сохраняют возможность превращений исключительно только в пределах своего тканевого типа. Под детерминированностью тканей, как и клеток, мы понимаем исторически сложившуюся их природу - характерные для них коренные свойства, наследственно закрепленные в процессе филогенетического развития.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'