Глава VII. Жизненный путь клетки в организме. Происхождение клеток и многоклеточности в эволюции
Каждая клетка проделывает определенный жизненный путь. Все клетки возникают в результате митотического деления материнской, но дальнейшая судьба их может быть различной. Одни из них на протяжении всей своей жизни не подвергаются существенной дифференцировке. Оставаясь в ткани ее камбиальными (стволовыми) малодифференцированными элементами, эти клетки вырастают до размеров материнской, а затем, в свою очередь, делятся. В период интерфазы, в данном случае аутосинтетической, они специфически мало или вовсе не функционируют, в основном подготавливаясь к предстоящему митотическому делению.
При систематически повторяющихся делениях камбиальных клеток, не выходящих из митотического цикла, смерти их с образованием трупа не наблюдается. Однако разделившись, данная клетка как какая-то индивидуальность исчезает. Так что срок жизни подобных клеток продолжается от деления до деления. Смерть их носит своеобразный характер - они, исчезая, продолжают в буквальном смысле слова жить материально в своих потомках.
Клетки со значительно выраженной дифференцировкой также могут вступить в митотический цикл, но этому должна предшествовать их дедифференцировка. Дедифференцируясь, данные клетки приобретают внешне сходство с эмбриональными. Они становятся резко базофильнее благодаря накоплению в цитоплазме массы рибосом, что свидетельствует о подготовке клеток к предстоящему усиленному синтезу белка. Ядерная оболочка образует многочисленные впячивания и выпячивания и таким путем увеличивается поверхность контакта кариоплазмы и цитоплазмы. Одновременно с этим в значительной мере претерпевают обратное развитие клеточные органеллы - митохондрии, пластинчатый комплекс, эндоплазматический ретикулум.
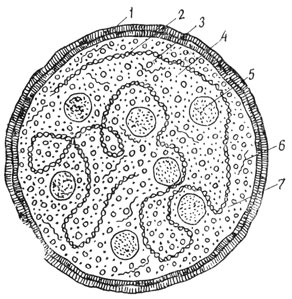
Рис. 31. Схема строения элементарного тела микоплазмы: 1 - липопротеидная мембрана; 2 - молекула ДНК; 3 - растворимый белок; 4 - метаболиты; 5 - рибосома; 6 - растворимая РНК; 7 - гиалоплазма
Наконец, имеется третья категория клеток, примером которых могут служить нервные клетки - невроны. Уже, будучи образованными к моменту рождения ребенка, большая часть их заканчивает свой жизненный цикл со смертью организма. Эти клетки достигают в своем развитии такой высоты дифференциации, что уже неспособны к дедифференцировке и митотическому делению.
Мы рассмотрели вопрос о жизненном цикле, то есть онтогенезе клеток. Коротко коснемся проблемы их филогенетического становления. Это совершенно не затронутая область, очень трудно поддающаяся исследованию. Соображения, которые высказываются на сей счет, основываются преимущественно на сравнительном материале, использовании данных о тонком строении наиболее примитивных из живых существ, а также на сопоставлении друг с другом различных компонентов клетки.
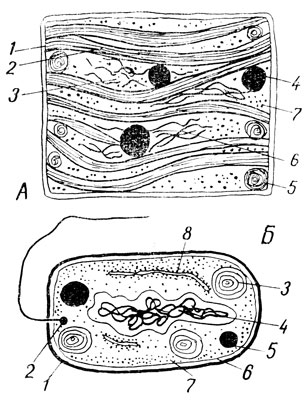
Рис. 32. А - предклетка сине-зеленой водоросли; 1 - плазмолемма; 2 - фотосинтетические пластинки; 3 - центроплазма; 4 - капля волютина (неорганического фосфора); 5 - пластинчатая система, эквивалентная митохондрии; 6 - нити ДНК; 7 - рибосомы; Б - бактерия: 1 - клеточная оболочка; 2 - жгутик; 3 - пластинчатая система; 4 - нуклеоид; 5 - капля волютина; 6 - плазмолемма; 7 - рибосомы; 8 - аналог шероховатого эндоплазматического ретикулума
Электронномикроскопическое изучение клетки, как мы видели, показало, что ее главнейшими структурными элементами являются мембраны, а затем гранулы и тончайшие нити. С этими же образованиями мы встречаемся и у одних из наиболее морфологически примитивно организованных живых существ - микоплазм. Микоплазмы относятся к числу самых мелких из известных микроорганизмов. Они имеют вид шариков диаметром около 0,15 мк и даже меньше, так что в оптическом микроскопе при самом сильном его увеличении они неразличимы. Встречаются как паразитические формы, с которыми связывают ряд заболеваний, так и свободно живущие. На схеме строения микоплазмы видно, что тело ее покрыто мембраной (рис. 31). Мембрана эта липопротеидная, подобно плазмолемме, и имеет примерно ту же толщину - около 100 Å. В теле микоплазмы располагаются нитевидная молекула ДНК типичного двуцепочечноспирального строения и некоторое количество рибосом. В ее протоплазме (которая еще не дифференцирована на цитоплазму и кариоплазму, поскольку отсутствует ядро) имеются различные ферменты, обеспечивающие протекание жизненных процессов, энергетического и пластического обмена. Микоплазма еще не клетка (вспомним, что по принятому определению клетка - это элементарная живая система, состоящая из цитоплазмы и ядра). В микоплазме не произошло обособления генетического аппарата в оформленное ядро, отграниченное оболочкой от цитоплазмы. Однако микоплазму можно рассматривать как одну из форм доклеточной организации живых существ, близкую той, которая дала начало клеткам на протяжении длительного исторического периода, измеряемого многими сотнями миллинов лет. При этом нужно иметь в виду, что, несмотря на свою морфологическую примитивность, микоплазмы не уступают клеткам в функциональном отношении. Их субмикроскопическая биохимическая организация, набор ферментов обеспечивают все жизненные процессы.
Следующую ступень в филогенетическом становлении клетки можно представить себе, проведя аналогию со строением сине-зеленых водорослей. Содержимое "предклеток" сине-зеленых водорослей дифференцировано на бесцветную центроплазму и фотосинтетические пластинки сине-зеленого цвета, образующие в совокупности хроматоплазму (рис. 32). В центроплазме залегают нити молекул ДНК толщиной 50 - 70 Å. В центроплазме имеются также рибосомы и эквиваленты митохондрий. Они имеют вид округлых пластинчатых образований и поэтому получили название интрацитоплазматических мембранных систем. Строятся они наружной мембраной "предклетки" - плазмолеммой. Кнаружи от этой мембраны имеется еще оболочка, а иногда и наружный чехол. Интересно, что встречаются нитчатые сине-зеленые водоросли, построенные из многих "предклеток" цилиндрической формы, заключенных в общую оболочку и разделенных перегородками. В этом случае пластинчатые образования хроматоплазмы могут, пронизывая перегородку, переходить из одной "предклетки" в другую.
В строении бактерий мы видим следующий шаг по пути от "предклетки" к клетке, причем среди этих микроорганизмов встречаются формы, находящиеся на разных этапах данного пути и уже приближающиеся по структуре, к клетке. Тело бактериальной "предклетки" подобно тому, что мы видели у сине-зеленых водорослей, одето липопротеидной мембраной - плазмолеммой, за которой располагается клеточная оболочка. Нити ДНК толщиной 40 - 60 Å либо собраны уже в пучки, свернутые в переплетающиеся жгуты толщиной примерно 0,2 мк, либо представлены одной очень длинной двойной нитью ДНК, образующей сложное переплетение. Эти структуры по своему функциональному значению соответствуют хромосомам. Однако оформленное ядро, одетое оболочкой, и у бактерий еще отсутствует. Хроматиновый аналог его у бактерий был назван нуклеоидом. Бактерии содержат, как и сине-зеленые водоросли, эквивалентные митохондриям, точнее кристам митохондрий, ламелярные системы, рибосомы (примерно такого же диаметра - 200 Å, как и в рассмотренных нами ранее клетках) и капли волютина (неорганического фосфата). Некоторые из внутрибактериальных мембран по своей функции аналогичны шероховатому эндоплазматическому ретикулуму. Бактерии движутся с помощью жгутика толщиной около 120 Å и длиной 3 - 12 мк. Жгутик образован тонкими нитевидными молекулами флателина (сократимого белка вроде миозина).
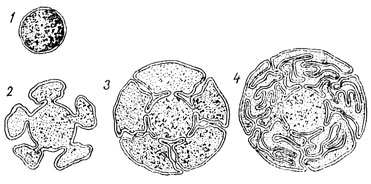
Рис. 33. Схема эволюции клетки по Робертсону: 1 - матричное вещество, окруженное элементарной мембраной; 2 - матричное вещество выпятилось, образовав псевдоподии; 3 - выпятившиеся мембраны впячиваются обратно, образуя парные ядерные мембраны и примитивный эндоплазматический ретикулум; 4 - дальнейшее развитие эндоплазматического ретикулума
Бактерии уже настолько близко подошли по своей организации к клеткам, что стали широко принятыми выражения "бактериальная клетка", "многоклеточные бактерии" (имеют в виду многие, так называемые микобактерии, в том числе и палочку Коха, вызывающую туберкулез), хотя все же они еще являются "предклетками", лишенными обособленного ядра.
На основании сравнительных исследований А. Н. Студитский высказал гипотезу об историческом развитии клеток из колоний бактериоподобных организмов, которые в процессе эволюции превратились в клеточные органоиды. Многие ученые в Советском Союзе и за рубежом предполагают в настоящее время, что митохондрии возникли из бактериальных организмов в результате их симбиоза с клетками. Основанием этому служат данные последних лет о сходстве митохондрий с бактериями по некоторым структурным и биохимическим свойствам. Это касается, в частности, липидного состава мембран и дыхательных ферментов, локализованных в них, нуклеотидного состава и физико-химических свойств ДНК, кольцевого строения ДНК и расположения его в центральной светлой зоне митохондрии, соответствующей нуклеоиду бактерий.
Робертсон предложил схему возможного исторического развития клеточных мембран (рис. 33). Их можно представить себе возникшими из элементарной перепонки, окружавшей каплю или комочек первичного живого тельца, претерпевшей постепенное усложнение в процессе филогенеза, вплоть до образования ядерной облочки, отделившей кариоплазму от цитоплазмы, и возникновения мембран эндоплазматического ретикулума и других органелл.
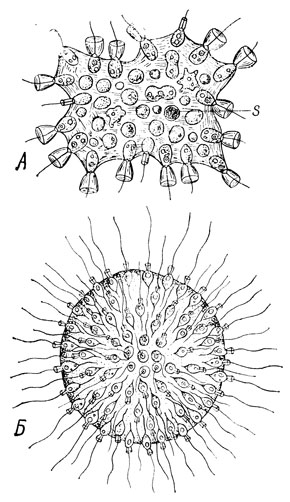
Рис. 34. А - колония Protospongia liaeckeli по Кенту. Б - колония воротничкового жгутиконосца Sphaeroecawolvox
Гораздо более разработанным является вопрос о происхождении в филогенезе из свободно живущих клеток многоклеточных организмов. Основная заслуга в этом принадлежит крупнейшему отечественному биологу И. И. Мечникову. В 1877 - 1886 годах он выступил с гипотезой колониального происхождения от жгутиконосцев - гетеротрофов (то есть с животным питанием) общего предка всех многоклеточных животных - первичного компактного многоклеточного организма, названного им паренхимеллой и им же позднее переименованного в фагоцителлу. По И. И. Мечникову, тело фагоцителлы построено из двух тканей-органов: наружного кинобласта и внутреннего - фагоцитобласта. Фагоцитобласт создавал внутреннюю среду организма и выполнял пищеварительную функцию, кинобласт отграничивал внутреннюю среду организма от наружной, осуществлял связь с этой средой, был одновременно и органом движения (за что и получил свое название). Постоянно происходило выселение клеток из кинобласта в фагоцитобласт.
Известный советский зоолог, лауреат Ленинской премии, профессор А. В. Иванов в монографии "Происхождение многоклеточных животных" (1968) отмечает важную роль Гаймана (1940, 1942) в дальнейшей разработке гипотезы И. И. Мечникова. В руководстве по зоологии беспозвоночных Гайман пишет, что Metazoa (многоклеточные животные) произошли от поляризованной полой сферической колонии жгутиконосцев, в которой сначала произошла дифференциация клеток на соматические и половые, а затем первые разделились на локомоторно-чувствительные и пищеварительные. Такая паренхимелла захватывала пищу клетками кинобласта, после чего эти клетки внедрялись в фагоцитобласт для пищеварения. Передний полюс тела данного животного был снабжен пучком чувствительных ресничек.
Существенное значение в подкреплении взглядов И. И. Мечникова о колониальном происхождении многоклеточных животных получила колония воротничковых жгутиконосцев Proterospongia, или Protospongia haeckeli, открытая Кентом (1880) в лондонском ботаническом саду (рис. 34А). Особи со жгутиками в этой колонии располагаются по периферии общей для всех клеток студенистой массы; другие клетки - амебовидные, лишенные жгутика и воротничка, погружены в нее. Находясь в поверхностном слое колонии, клетки захватывают пищевые частицы, а нагрузившись ими, сбрасывают жгут и воротничок и погружаются внутрь колонии, где переваривают фагоцитированные частицы. После этого они вновь направляются на периферию, вдвигаются между находящимися там жгутиконосцами, образуют на своем свободном конце жгут и воротничок и принимаются за фагоцитирование новых пищевых частиц. Клетки периферического и внутреннего слоев колонии оказываются дифференцированными по-разному, но эта дифференцировка обратимая, лабильная. Она носит функциональный характер и не закреплена. Таким образом, периферический слой протероспонгии может служить прообразом кинобласта фагоцителлы Мечникова, а внутренний слой - фагоцитобласта.
В связи со сказанным следует сослаться на исследование А. А. Захваткина (1949), показавшего, что с увеличением количества клеток у колониальных жгутиковых колонии становятся все более интегрированными. Так, в колониях с числом клеток 4 - 8 интеграция полностью отсутствует и все клетки - зооиды равноценны. При распаде колонии каждый зооид образует новую колонию бесполым путем или превращаясь в половую клетку-гамету. Все гаметы одинаковы. В колониях из 16 клеток интеграция лишь зачаточная и они мало устойчивы. Наблюдаются незначительные различия между гаметами. Колонии из 32 - 128 зооидов устойчивы. Часть передних клеток (у 32-клеточной колонии) или все клетки передней половины колонии (у 64 - 128-клеточной) теряют способность к размножению. Гаметы резко отличаются друг от друга.
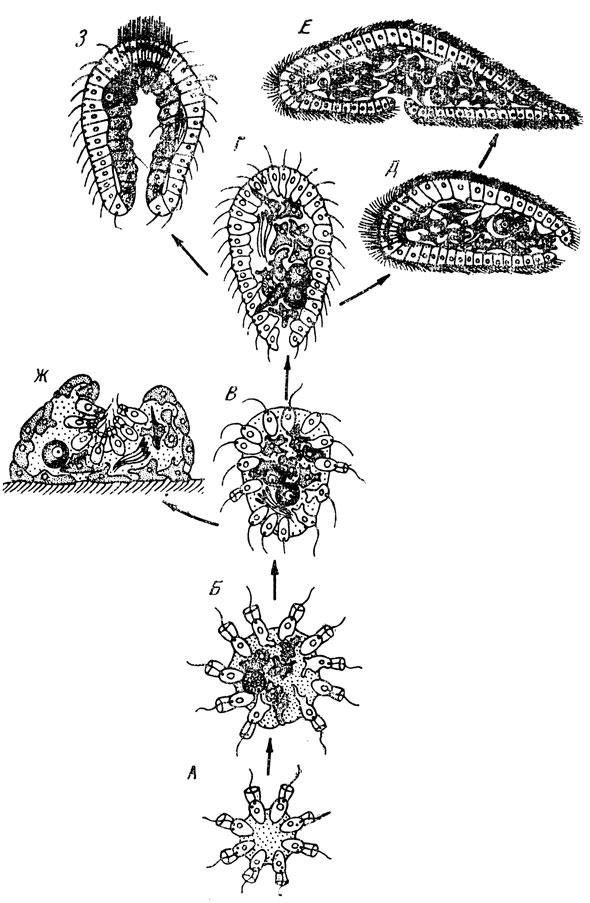
Рис. 35. Главнейшие предполагаемые стадии филогенеза многоклеточных животных (по А. В. Иванову): А - колония типа Sphaeroeca Б - колония типа Protospongia; В - ранняя фагоцителла; Г - поздняя фагоцителла; Д - первичная турбеллярия - появление рта и билатеральной симметрии; Е - примитивная бескишечная турбеллярия - усиление дифференциации клеток, смещение рта на брюшную сторону; Ж - примитивная губка - переход к сидячему образу жизни, замена локомоторной функции киноцитов гидрокинетической; 3 - первичное кишечнополостное - образование рта, эпигелизация фагоцитобласта
Колонии из 512 и большего числа зооидов, такие, как Вольвокс, уже близки к многоклеточному организму. Основная масса клеток - соматические. Они расположены в пласт, наподобие эпителия, и образуют однослойную стенку шарообразной колонии, полость которой заполнена слизью (рис. 34Б). Друг с другом зооиды соединены тонкими тяжами цитоплазмы. Высокая степень интеграции такой колонии проявляется в дифференциации переднего и заднего полюсов, в согласованной работе жгутиков и упорядоченном движении колонии. Половые клетки немногочисленны и представлены уже спермиями и яйцеклетками. Сам А. А. Захваткин пришел к заключению, что колонии Вольвокса, состоящие из тысяч зооидов, должны рассматриваться как примитивные многоклеточные животные.
Крупнейший специалист по данной проблеме А. В. Иванов считает, что наиболее приемлемой гипотезой происхождения Metazoa можно признать только концепцию фагоцителлы И. И. Мечникова. На основании своих совместных с О. М. Ивановой-Казас исследований (1967) он высказал мнение, что исходные колонии, от которых произошли предки Metazoa, мало отличались от современных воротничковых жгутиконосцев, таких, как Sphaeroeca.
В упомянутой монографии А. В. Иванов намечает наиболее вероятные стадии ранней филогении Metazoe (рис. 35). Видимо, первым шагом на пути становления многоклеточных животных было появление монотомической колонии типа Sphaeroeca (монотомия - простое деление клетки надвое, оно часто встречается у Protozoa и типично для тканевых клеток Metazoa), то есть свободноплавающая, шаровидная, с периферическим расположением жгутиконосцев. Все особи колонии однородны. Размножение только бесполое - путем распада колоний на отдельные клетки. Схематически такого типа колония показана на рисунке 35А.
Следующий шаг на пути к Metazoa представляла колония со значительной общей интеграцией (рис. 35Б). Появление полового процесса и палинтомического характера начальных стадий онтогенеза (палинтомия - деление, повторяющееся многократно и не сопровождающееся ростом дочерних клеток; примером может служить дробление зиготы у Metazoa, оно присуще многим Protozoa). Первая дифференциация клеток на соматические и половые. Удлинение палинтомического отрезка жизненного цикла и превращение его в период дробления. Дифференциация переднезадней оси тела и появление радиальной симметрии неопределенно большого числа. Развитие анизогамии (различий между гаметами). Некоторое приближение к этой филогенетической стадии представляет собой колония типа протероспонгии.
Наконец, возник свободноплавающий организм типа мечниковской фагоцителлы (рис. 35В). Сначала лабильное, потом стойкое разделение клеток на наружный кинобласт и внутренний фагоцитобласт, как представлял себе это И. И. Мечников. Разделение всего онтогенеза на эмбриональный и постэмбриональный периоды.
Исторически сложившийся таким образом плавающий организм, очень близкий к фагоцителле, вероятно, сохранял еще первичную способность к бесполому размножению. Оно, возможно, осуществлялось на первых порах распадением тела на отдельные клетки, которые давали начало многоклеточным особям. Однако такая форма бесполого размножения, совершенно несвойственная Metazoa, затем, в связи с усилением дифференциации и интеграции, уступила место разделению тела на более крупные многоклеточные фрагменты (в состав их входили и кинобласт, и фагоцитобласт), которые путем незначительной регуляции формы превращались в новые индивиды.
Дальнейшую эволюцию можно представить в соответствии с идеями И. И. Мечникова следующим образом. Внутренние амебоидные клетки фагоцитобласта, вероятно, могли заглатывать более крупные пищевые частицы, чем киноциты. На более поздних этапах эволюции, когда сформировалась уже поляризованная свободноплавающая фагоцителла, наиболее удобным местом для захвата пищи оказался задний полюс тела, где вследствие биения жгутов кинобласта образуется мертвое пространство, в котором собираются пищевые частицы. Естественно предположить поэтому, что именно здесь, на физиологически заднем конце тела, путем расхождения киноцитов у поздней фагоцителлы появилось первичное ротовое отверстие, через которое фагоциты могли захватывать пищу (рис. 35Г).
Из ныне обитающих на Земле животных ближе всего стоят к фагоцителле губки и бескишечные ресничные черви Acoela (рис. 35Д и Ж). От фагоцителлы произошли и общие предки кишечнополостных - сидячих (Cnidaria) и плавающих (Ctenophora). Их фагоцитобласт поляризовался, эпителизировался целиком.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'