Глава VI. Наследственная передача признаков и клеточная дифференцировка
Хромосомная теория наследственности получила в последние годы благодаря работам биохимиков кардинальное подкрепление. Однако и сегодня в центре внимания исследователей находится вопрос о существовании внехромосомной наследственности и ее удельном весе в генетической передаче признаков. Многочисленные факты свидетельствуют о том, что решающая роль в этом процессе принадлежит ядру. Последнее можно проиллюстрировать исключительно демонстративными экспериментами академика Б. Л. Астаурова. Они были проведены на половых клетках двух видов тутового шелкопряда.
От дикого вида шелкопряда брали яйцеклетки и оплодотворяли их сперматозоидами шелкопряда культурного вида. Перед оплодотворением ядро яйцеклетки подвергали разрушению путем рентгеновского облучения. У тутового шелкопряда при оплодотворении в яйцеклетку проникают головки нескольких сперматозоидов. В данных опытах, когда яйцеклетка оказывалась безъядерной, ядро зиготы образовывалось в результате слияния двух мужских ядер, так что клетки развивающейся личинки были диплоидными. Сама же личинка обладает, как было обнаружено во всех таких экспериментах, исключительно признаками шелкопряда культурного вида, то есть целиком обусловленными ядром.
А как же обстоит дело с потенциями ядер дифференцирующихся клеток многоклеточных организмов? Сохраняют ли ядра соматических клеток способность развития целого организма?
Мы уже отмечали, что все соматические клетки, возникающие путем многократных митозов из зиготы, обладают таким же набором хромосом и, следовательно, имеют совершенно такой же набор генов. Уже давно было известно, что каждая из двух дочерних клеток зиготы может дать, развившись, полноценный организм. Для этого нужно лишь разъединить их друг от друга, перетянув шнурком. Такие два существа, возникшие из одной оплодотворенной яйцеклетки, называют идентичными, или однояйцевыми близнецами. Они довольно часто встречаются и среди людей. В отличие от неидентичных, разнояйцевых близнецов, они обладают тождественным геномом (всей совокупностью наследственных зачатков - генов), заложенным в их хромосомах, которые, таким образом, ничем не отличаются друг от друга. Поэтому однояйцевые близнецы всегда одного пола и по внешнему виду трудно различимы. Тождественны у них и носители жизни - клеточные белки. Их не может различить даже такой тончайший анализатор как система иммуногенеза организма.
Если ввести во внутреннюю среду человека чужеродные белки либо нуклеиновые кислоты или сложные углеводы полисахариды), они будут действовать как антигены - на их начнут вырабатываться соответствующими клетками ткани внутренней среды организма особые белки - антитела. Антитела строго специфичны и вырабатываются специально на каждый отдельный антиген. Антитела обладают свойством, взаимодействуя с соответствующими антигенами, способствовать выведению их из организма. Данная защитная реакция носит название иммунной. На ней основан иммунитет, то есть невосприимчивость организма к определенным антигенам, на которые его специальными клетками (лимфоидными элементами и в первую очередь плазматическими) были уже выработаны антитела.
Лучшим показателем тождественности белков у однояйцевых близнецов являются результаты пересадки тканей и органов от одного организма другому. Если пересадить, например, лоскут кожи от одного человека другому, даже его разнояйцевому близнецу, он неминуемо будет отторгнут вследствие разрушающего действия антител. Пересадки от однояйцевого близнеца удаются так же хорошо, как и от самого себя.
Генетическая тождественность однояйцевых близнецов и такие же различия по этому признаку между разнояйцевыми близнецами, как и между обычными братьями и сестрами, дают возможность, сравнивая развитие тех и других, изучать роль наследственных факторов и среды в возникновении разных особенностей организма и болезней. Факторы среды могут вызывать у развивающихся в разных условиях жизни однояйцевых близнецов различия физического характера, и особенно в высшей нервной деятельности. В отношении заболеваний вопрос об их наследственности в данном случае решается положительно, если сходство по этому признаку между однояйцевыми близнецами окажется достоверно большим, чем между разнояйцевыми.
Опыты, проведенные на амфибиях, показали, что не только дочерние клетки зиготы могут дать начало целому организму. Шпеманом (1928) был поставлен следующий опыт. Он осторожно перетягивал шнурком зиготу так, чтобы ядро находилось в одной из ее половин. Это не препятствовало дроблению в той половине, где находилось ядро, а когда оно достигало стадии 16 бластомеров (так называются молодые эмбриональные клетки), то есть после четырех делений, он отпускал петлю шнурка настолько, чтобы в безъядерную половину зиготы могло проникнуть одно из 16 ядер ее раздробившейся второй половины. Затем петлю снова затягивал. В результате из одной зиготы развивалось два организма, только второй зародыш несколько запаздывал.
Бриис и Кинг (1955) разработали методику пересадки в энуклеированное (лишенное ядра) яйцо лягушки ядра какой-нибудь "3 эмбриональных клеток головастика. Известно, что яйца лягушек способны к партеногенетическому (девственному) развитию после активирования их уколом иглы. Этой операцией и начинается данная методика трансплантации ядер. Затем активированное яйцо энуклеируют с помощью стеклянной пипетки и вносят в него стеклянной микропипеткой нужное ядро, засосанное из выбранной клетки-донора. Указанными авторами и другими было установлено, что даже при пересадке в энуклеированные яйца клеточных ядер от зародышей поздних стадий развития частично развиваются совершенно нормальные головастики.
Было получено развитие нормальных головастиков при пересадке в энуклеированные яйца ядер, взятых из уже дифференцированных клеток эпителия кишки или почки головастика. Оказалось, что при трансплантации ядер из кишечной выстилки головастика шпорцевой лягушки могут быть получены нормальные взрослые животные, но только в 2% сделанных пересадок. Гибель части яиц с пересаженными ядрами, а также дефекты в их развитии можно объяснить повреждающими воздействиями, испытываемыми ядрами, во-первых, от манипуляций с ними при пересадке, во-вторых, от окружающей цитоплазмы уже после трансплантации. Относительное увеличение количества неудач с повышением уровня развития зародышей, от которых берутся ядра для трансплантации, возможно, зависит от усиливающейся по мере дифференциации клеток, повреждаемости ядер.
Детальное исследование показало, что имеется прямая зависимость между состоянием хромосомного набора в клетках развивающихся зародышей и характером развития. В тех случаях, когда развитие быстро прекращалось, в клеточных ядрах обнаруживались различные хромосомные нарушения. Когда же развивалась личинка, пусть даже с теми или другими дефектами, хромосомные нарушения в клетках были менее выражены. В клетках нормально развивающихся головастиков никаких нарушений в хромосомном наборе не наблюдалось.
Из сказанного следует с очевидностью, что генный набор, содержащийся в ядрах дифференцированных соматических клеток, может, конечно, при соответствующих условиях обеспечить, подобно зиготе, нормальное развитие целого организма. К этому следует добавить, что при пересадке ядер в энуклеированные яйцеклетки, в случае использования половых клеток от представителей разных подвидов или разновидностей, внешний вид взрослой особи всегда соответствовал отцовской форме, то есть определялся исключительно ядром.
Примерно полвека назад был широко распространен взгляд, что генами, наследственными зачатками, заложенными в хромосомах, определяются признаки, которыми отличаются друг от друга лишь представители разновидностей и пород, а за более существенные признаки, характеризующие виды, роды, семейства и еще более высокие таксономические единицы, отвечают наследственные зачатки цитоплазмы, объединяемые общим названием плазмой.
В опытах со взаимным обменом ядер у амеб A. proteus и A. discoides было обнаружено, что форма тела определяется цитоплазмой. У A. proteus псевдоподии немногочисленны, а у A. discoides их много, и тело имеет зубчатые очертания. Сохранение внешних признаков того вида амебы, от которого была взята цитоплазма при трансплантации, наблюдалось на протяжении всех исследованных 600 поколений. Интересно, что при этом выявилось закономерное влияние цитоплазмы на размеры ядра. У A. proteus оно имеет диаметр 45 мк, у A. discoides - 38 мк. После реципрокной пересадки ядер размеры их становились в среднем такими же, как у вида, которому принадлежала цитоплазма. Аналогичным образом было показано влияние цитоплазмы на ядро в опытах с пересадкой ядер в энуклеированные яйца лягушек другого вида (от Rana pipiens к R. sylvaticum).
Мы уже говорили, что в целом ряде органоидов цитоплазмы доказано наличие ДНК. В отношении митохондрий установлено, что меченый структурный компонент ДНК тимидин включается в митохондрии в 10 раз активнее, чем в ядре.
Поскольку известные уже плазмогены представлены молекулами ДНК, как и гены ядра, можно думать, что и механизм их действия сходный. Заложенная в плазмогенах органоидов наследственная информация должна обеспечивать их репродукцию. Считают, что действие плазмогенов находится все же под контролем хромосомных генов.
Как же сейчас, исходя из данных современных цитологических и молекулярнобиологических исследований, следует рассматривать вопрос о детерминации клеток и путях их дифференциации? Чем обусловлено, что клетки, возникающие из одной и той же оплодотворенной яйцеклетки, проделывают разный путь развития и превращаются в не сходные по своему виду и свойствам клеточные элементы разных тканей?
Говоря в одной из предыдущих глав о клеточном митотическом цикле, мы отметили, что клетки могут выходить из него и вступать в гетеросинтетическую интерфазу В это время и происходит их дифференцировка, то есть взаимосвязанная функциональная и морфологическая специализация. О том, как реализуется в дифференцирующихся клетках заложенная в ядре генетическая информация, помогли разобраться наблюдения над гигантскими политенными хромосомами слюнных желез развивающихся двукрылых.
Об этих хромосомах мы уже говорили. Добавим только, что длина их может достигать 400 мк. Они могут быть построены из тысяч хромонем и все же однозначные точки (хромомеры) всех хромонем лежат на одном уровне (рис. 6). Поэтому в гигантской политенной хромосоме легко выявляется специфическое для каждой из них последовательное расположение дисков (поперечных полос) и промежутков между ними. Подобных дисков насчитывают в некоторых политенных хромосомах до 6000. Каждый такой диск отвечает, как было установлено, определенному гену или группе генов. Так что в структуре политенной хромосомы отчетливо выражено линейное расположение генов.
В процессе развития личинок этих насекомых указанные диски хромосом образуют вздутия, получившие наименование пуфов (рис. 29). Пуфы появляются в разное время в различных участках гигантских хромосом. Пеллинг показал, что образование пуфов обусловлено синтезом РНК. Так возникла вполне обоснованная мысль, что пуфы представляют собой дерепрессированные гены, в то время как в остальных участках хромосом гены остаются в репрессированном состоянии. Сейчас считают, что хромосома в диске, не образующем пуфа, обладает ДНК, а точнее нуклеогистоном, уплотненным и сильно спирализованным. Пуф возникает в результате разрыхления в данном участке нуклеогистона. В результате раскручивания вторичных спиралей возникают "шлейфы", образующие пуф. В таких шлейфах и синтезируется и-РНК, в свою очередь определяющая синтез соответствующих белков. Данный участок хромосомы возвращается позднее в репрессированное состояние вследствие уплотнения и спирализации молекулы ДНК, связанной с гистоном.
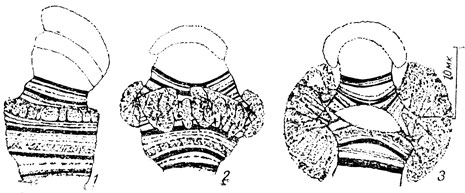
Рис. 29. 1 - 3 - последовательные стадии развития пуфа на одном из участков гигантской хромосомы (из Богена)
Крёгер изменял среду, окружавшую гигантские хромосомы с выраженными пуфами, перенося клеточные ядра из слюнных желез уже развитых личинок в цитоплазму начавших развитие яиц. В результате вздутия некоторых хромомеров исчезали, зато появлялись пуфы на других. В зависимости от стадии развития яиц их цитоплазма вызывала появление пуфов в разных местах хромосом. В опытах Гёрдена, проведенных на шпорцевой лягушке, было установлено, что при пересадке ядер в цитоплазму энуклеированных яйцеклеток, находящихся на разных стадиях развития, синтез ДНК и разных видов РНК регулируется цитоплазмой. Пересаженные ядра синтезируют именно те ДНК и РНК, которые должны были продуцировать собственные ядра подопытных яйцеклеток на данной стадии развития. Гёрден отмечает набухание трансплантированных ядер в результате поступления в них белков цитоплазмы и рассматривает это как способ регулирования цитоплазмой активности ядра.
Из приведенных экспериментов был сделан правильный вывод, что дифференцировка - это результат ядерно-цитоплазменного взаимодействия. В процессе дифференцировки генетические свойства ядер проявляются под влиянием цитоплазматических факторов. С другой стороны, дифференцировку клеток нельзя представлять себе как обусловленную исключительно одной генетической информацией. Она реализуется на основе взаимосвязи процессов разного порядка, причем весьма важное значение имеют межклеточные взаимодействия, в частности, степень контакта клеток и их взаиморасположение в пространстве. Клетки обмениваются информацией, взаимодействуя друг с другом и непосредственно своими поверхностями, и химическим путем.
Приобретаемые на ранних стадиях онтогенеза клетками определенных эмбриональных закладок и формирующихся тканей общие для них и стабильные характерные особенности в расположении на хромосомах участков дерепрессии и последовательности их включения во времени представляют собой проявления так называемой эпигеномной наследственности. Эпигеномная наследственность определяется цитоплазматически обусловленным частичным блокированием или деблокированием наследственной информации, заложенной в клеточных ядрах.
Академик А. А. Заварзин всегда подчеркивал, что клеток "вообще" не бывает. Всякая соматическая клетка является структурным элементом системы более высокого порядка-ткани. Поэтому клеточная дифференцировка, по существу, является тканевой. Клетки, объединенные общностью проявлений эпигеномной наследственности, и образуют в совокупности ту или иную ткань. Известный советский гистолог профессор В. П. Михайлов определяет ткань как "частную систему организма, состоящую из клеток с одинаковым образом реализуемой эпигеномной наследственностью".
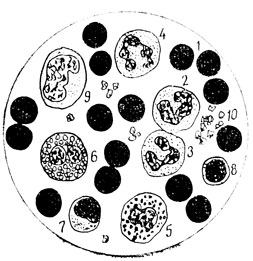
Рис. 30. Мазок крови человека: 1 - эритроциты; 2 - палочко-ядерный нейтрофил; 3 - молодой нейтрофил; 4 - зрелый нейтрофил; 5 - базофил; 6 - эозинофил; 7 - средний лимфоцит; 8 - малый лимфоцит; 9 - моноцит; 10 - кровяные пластинки (тромбоциты)
Говоря о клеточной дифференцировке в общем виде, мы фактически рассматривали вопрос о приобретаемой всеми соматическими клетками в процессе индивидуального развития организма определенной детерминированности - закрепленности, отражающей их тканевую принадлежность. Тканевая детерминированность, согласно представлениям школ А. А. Заварзина и Н. Г. Хлопина, это эволюционно обусловленная, возникающая и закрепляемая в каждом онтогенезе тканевая специфичность. Все клетки одного тканевого типа обладают общими свойствами и потенциями, качественно отличающими их от клеток других тканевых типов. Их конкретная дифференцировка, связанная с морфофункциональной специализацией как структурных элементов той или иной тканевой системы, входящей в состав разных органов, может быть весьма различной. Однако во всех своих функциональных проявлениях и дифференцировках каждая соматическая клетка ведет себя в соответствии с природой ткани, структурным элементом которой она является. Эта, так сказать, частная дифференцировка клеток определяется конкретными местными условиями, всякого рода коррелятивными взаимоотношениями с другими клеточными элементами данной ткани и регулируется функциональными потребностями органа и организма в целом. Так, например, возникая у взрослых особей из камбиальных (мало дифференцированных клеток) гемоцитобластов, детерминированных как кроветворные клеточные элементы, клетки крови проделывают различную частную дифференцировку. Для примера можно привести рисунок мазка крови человека, на котором видны безъядерные эритроциты (красные кровяные клетки, разносящие кислород по организму) и пять видов лейкоцитов (белых кровяных клеток, выполняющих в основном защитную функцию): зернистые - нейтрофилы, эозинофил, базофил и незернистые - моноцит, средний и малый лимфоцит (рис. 30). Процесс дифференцировки интенсивно новообразующихся в кроветворных органах форменных элементов, то есть клеток крови, настолько точно регулируется, что наблюдающиеся при разного рода заболеваниях отклонения в процентном содержании тех или иных клеток крови от нормы весьма помогают в установлении диагноза и определении тяжести болезни. Вот почему врачи так широко прибегают к анализу крови.
Однако передифференцировка клеток в пределах тканевого типа обычное явление. Она сказывается прежде всего в некотором специфическом изменении синтезируемых белков, но детерминация клеток, носителей определенной тканевой наследственности, природа их остается неизменной. Возвращаясь к тому же примеру - форменным элементам крови, мы можем указать на часто наблюдающуюся в культурах ткани и в экспериментах на целых организмах передифференцировку лимфоцитов в зернистые лейкоциты и даже в клетки - волокнообразователи соединительной ткани - фибробласты.
Совершенно иначе обстоит дело с метаплазией (по-гречески "метаплассеин" - преобразовывать) клеток одного тканевого типа в другой. В отношении млекопитающих и человека можно уверенно сказать, что такие тканевые преобразования не происходят. По-видимому, все другие участки хромосом, кроме отвечающих клеточным элементам данного тканевого типа, оказываются прочно заблокированными, и дерепрессия в них становится поэтому невозможной. Ядра клеток различных тканей тела не различаются ни по составу, ни по содержанию ДНК, однако в разных тканях в активное состояние переходят различные участки хромосом.
Остановимся коротко на опытах, в которых с исключительной наглядностью выступает тканевая природа у детерминированных клеток. Мы имеем в виду эксперименты по объединению специально разобщенных клеток. Было известно, что при беспорядочном смешении в одном сосуде с морской водой разъединенных "клеток тела губок разного вида, отличающихся по цвету, наблюдается собирание вместе клеток одного цвета, то есть по видовому признаку. Вероятно, это связано с поступлением в воду каких-то видоспецифических веществ, вырабатываемых каждой клеткой, и способностью клеток воспринимать эти сигналы и реагировать на них.
Позднее Москона обнаружил, что в смешанных культурах ткани разобщенные клетки зародышей мыши и курицы, подобно клеткам разных видов губок, объединяются в скоплении в соответствии со своей видовой принадлежностью. Однако когда для экспериментов были взяты разобщенные клетки разных тканей, то объединение их происходило не по видовому, а по тканевому признаку. В смеси клеток, среди которых имелись почечные клетки зародышей мыши и курицы, образовывались почечные канальцы, состоящие из клеток, принадлежащих животным обоих видов. Таким образом, тканеспецифические факторы, благодаря которым клетки распознают друг друга и объединяются в тканевую систему, оказываются более действенными, чем видоспецифические.
Во многих случаях в качестве факторов, контролирующих активность генов, доказано действие гормонов - продуктов, вырабатываемых железами внутренней секреции. Эти вещества специфического действия поступают из желез в кровь и разносятся ко всем тканям и клеткам тела. Под влиянием гормонов в тех или иных клетках высших животных и человека включаются или выключаются отдельные гены или группы их, что приводит к синтезу соответствующих белков - структурных или ферментных и может в известных пределах (внутри тканевого типа) вызывать переход клеток на новый путь развития.
Дифференцировка клеток выражается прежде всего в синтезе специфических для каждой из них белков. В целом же дифференцировка клетки является конечным отражением действия на нее регулирующих механизмов, решающих, какие именно из генетических локусов и в какой момент становятся активными. Под влиянием этих координирующих воздействий (исторически обусловленных и закрепленных в онтогенезе) каждая клетка в процессе индивидуального развития организма снижает количество возможных состояний до небольшого числа реально достигаемых, и эти состояния стабилизируются настолько, что колебания условий среды и метаболические ошибки не вызывают перехода клеток одного тканевого типа в другой (например, печеночных в соединительнотканые).
Развитие организма - запрограммированный процесс, который реализуется на основе филогенетически обусловленной и закрепленной естественным отбором определенной последовательности в репрессии и дерепрессии отдельных структурных элементов генома. В процессе развития тканей и органов в результате их взаимодействия возникают индуцирующие влияния. Боннер (1966) указывает, что индукция - это наведение на какой-то определенный путь развития. Эмбриогенез в целом и в отдельных его частях контролируется сложной сетью таких индукций - "переключений", координированных в своей деятельности. Цитоплазматически обусловленное частичное блокирование и деблокирование генетической информации определяет эпигеномную наследственность "леток и образуемых ими тканей.
О влиянии окружающей среды на генетическую активность ДНК мы находим интересное высказывание у виднейшего американского молекулярного биолога-биохимика Мирского. В своем заключительном слове председателя на симпозиуме, проходившем в Лондоне в начале декабря 1965 года в честь нашего крупнейшего ученого академика В. А. Энгельгардта и посвященном проблеме гистонов, Мирокий подчеркнул, что биология в настоящее время вышла за рамки абстрактных и жестких ограничений классической генетики и теперь становится ясным, что хромосомы, подобно другим центрам жизненной активности, являются объектом регулирования извне по принципу обратной связи.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'