Глава V. Половые клетки и цитологические основы некоторых наследственных заболеваний
Мы говорили, что соматические клетки содержат двойной, диплоидный набор хромосом, а зрелые половые - одинарный, гаплоидный. Диплоидный набор хромосом имеется и в еще не созревших половых клетках. Уменьшение вдвое количества хромосом и соответственно ДНК, обозначаемое как их редукция, происходит в процессе гаметогенеза, то есть развития половых клеток - гамет. Редукция вдвое числа хромосом в гаметах является подготовкой к будущему оплодотворению, при котором восстанавливается диплоидный их набор, благодаря объединению гаплоидных хромосомных наборов сперматозоида и яйцеклетки.
Гаметогенез протекает в половых железах: в семенниках у мужского организма и в яичниках - у женского. Соответственно он получил название сперматогенеза и оогенеза. Общими для спермато- и оогенеза являются 3 первых периода гаметогенеза: размножение, рост и созревание. Развивающиеся мужские половые клетки проходят дополнительно еще четвертый период - формирование (рис. 26).
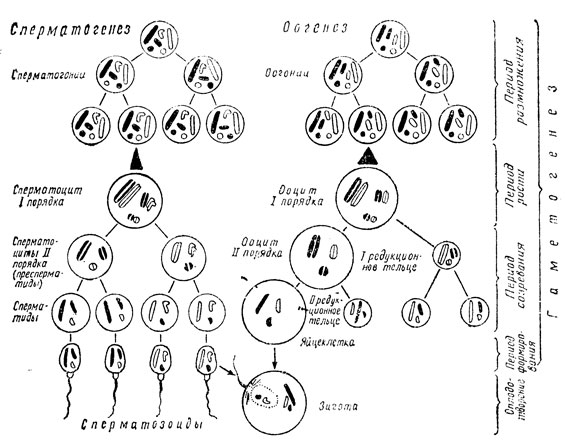
Рис. 26. Схема гаметогенеза (из А. Г. Кнорре)
В первом периоде гаметогенеза половые клетки усиленно делятся митотически и количество их нарастает. В данном периоде половые клетки носят название соответственно спермато- и оогониев. На схеме для простоты показан случай, когда гаплоидный набор включает всего 3 хромосомы, причем одна из них половая - гоносома, или гетерохромосома (по-гречески "гетерос" означает другой) и две неполовые - аутосомы. В клетках один гаплоидный набор зачернен, второй дан контурными линиями. Гомологичные, однозначные аутосомы обоих наборов нарисованы одинаковой формы и размеров (это наиболее длинная черточка и кружок). Гетерохромосомы изображены неодинаково - прямой черточкой (X-хромосома) и такой же длины изогнутой черточкой (Y-хромосома). Хромосомы лежат в клетке обособленно.
Перешедшие в период роста половые клетки обозначаются как спермато- и ооциты I порядка. Они увеличиваются в размерах, особенно резко ооциты, и их ядерный аппарат претерпевает перестройку. Гомологичные хромосомы ложатся параллельно друг другу, образуя биваленты, число которых равно количеству хромосом в гаплоидном наборе. Каждая из двух хромосом бивалента в свою очередь является парной структурой - диадой, поскольку состоит из двух сестринских хроматид. Когда щель между этими хроматидами становится хорошо выраженной, биваленты выглядят уже как тетрады. Количество тетрад отвечает гаплоидному числу хромосом. Общее же число выявляемых хроматид - будущих хромосом зрелых половых клеток является тетраплоидным. Тетраплоидно и количество ДНК в спермато- и ооцитах перед периодом созревания.
Затем наступает период созревания, который характеризуется мейозом ("мейозис" означает уменьшение) - двумя быстро следующими друг за другом делениями половых клеток, при котором и происходит редукция хромосом. Мужские половые клетки, проделавшие первое деление созревания, называются сперматоцитами II "порядка, или пресперматидами, а соответствующие им женские половые клетки - ооцитами II порядка. После второго деления созревания пресперматиды становятся оперматидами, ооциты II порядка - зрелыми яйцеклетками. Первое деление созревания является редукционным. При этом делении между дочерними клетками распределяются целые хромосомы - диады и хромосомный набор становится гаплоидным. Второе деление созревания называют уравнительным, эквационным, так как между дочерними клетками расходятся половинки хромосом (диад), по существу их хроматиды.
Интерфаза между первым и вторым делением созревания может быть совсем короткой или даже вообще отсутствовать, так как здесь в это время не происходит в клетках ни редупликации ДНК, ни удвоения числа хромосом. Снова наступает деление, теперь уже двух клеток, и каждая из внучатых клеток получает по одной хромосоме из диады. Таким образом, в каждую из этих четырех клеток, возникших в результате 2 делений созревания, попадает по одному из элементов тетрады. Гаплоидному числу тетрад отвечает гаплоидное число в хромосомном наборе клеток, проделавших мейоз. Точно так же тетраплоидное в начале мейоза количество ДНК (в ооците и сперматоците I порядка) после разделения на 4: части становится гаплоидным в конце его.
При сперматогенезе из каждого сперматогония, вступившего в период роста, получаются в итоге делений созревания 4 полноценные сперматиды. При оогенезе возникающий из ооцита I порядка ооцит II порядка сохраняет почти полностью размеры материнской клетки. Вторая же дочерняя клетка получает половину хромосомального материала материнской и лишь ничтожную часть ее цитоплазмы. Эта мелкая клетка получила наименование редукционного тельца. Аналогичная картина повторяется и при втором делении созревания - ооцит II порядка дает начало яйцеклетке примерно равного с ним размера и второму редукционному тельцу. В это же время делится надвое и первое редукционное тельце. В итоге из одного оогония, перешедшего из периода размножения в период роста, а затем в период созревания, образуется только одна зрелая яйцеклетка. Это является выгодным для продолжения рода приспособлением - в яйце остается весь накопленный ооцитом I порядка в периоде роста запас питательных веществ, необходимых для обеспечения начальных этапов развития будущего эмбриона.
В периоде формирования происходит перестройка сперматиды типичной клетки в сперматозоида, имеющего очень сложное строение, обеспечивающее его роль как активного, подвижного партнера в акте оплодотворения. В этой перестройке участвуют все составные части сперматиды.
Прежде всего ее центриолы располагаются одна за другой, определяя таким образом длинную ось будущего сперматозоида (рис. 27А). Среди многих мелких проакросомных гранул, образовавшихся еще в сперматоците - в центре его аппарата Гольджи, который движется теперь к переднему концу клетки (рис. 27Б-3), появляется одна крупная акросомная ("акрон" означает верхушка) гранула, которая ложится затем у ядра в месте его будущего головного полюса (рис. 27В-9). Пластинчатый комплекс при этом редуцируется, давая начало пузырьку-акробласту, охватывающему акросомальную гранулу. Тело сперматиды начинает постепенно удлиняться, а ядро клетки все более и более уплотняется. Оно располагается у переднего конца формирующегося сперматозоида. Проксимальная центриола ложится сзади от ядра, а дистальная образует, подобно кинетосоме, жгутик. Потом она делится на две части, причем задняя из них принимает форму колечка (рис. 27В-8) и отодвигается от передней, скользя по разрастающемуся жгутику - будущей осевой нити хвостика сперматозоида. Колечко задерживается у заднего края клетки. К этому времени митохондрии оказываются уже большей частью скопившимися у осевой нити. Акробласт, разрастаясь, надвигается в виде колпачка на переднюю часть ядра.
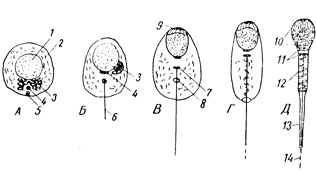
Рис. 27. А - Д - схема последовательных стадий формирования сперматозоида: 1 - ядро сперматиды; 2 - митохондрии; 3 - внутриклеточный сетчатый аппарат (пластинчатый комплекс); 4 - проксимальная центриола; 5 - дистальная центриола; 6 - жгутик; 7 - передняя часть дистальной центриолы; 8 - задняя часть дистальной центриолы; 9 - акробласт; 10 - головка сперматозоида; 11 - шейка; 12 - промежуточный отдел; 13 - главный отдел хвостика; 14 - концевой отдел хвостика
В конце периода формирования в сперматозоиде хорошо выражена дифференцировка на отделы: головку, представляющую собой в основном уплощенное и очень компактное ядро, одетое спереди чехликом, под которым у самого края лежит акросома; шейку, образованную центриолами; промежуточный, связующий отдел и хвостик. В промежуточном отделе сконцентрированы все митохондрии, которые охватывают по спирали осевую нить. Дистальной границей промежуточного отдела служит замыкающее центриольное кольцо. В цитоплазме здесь имеется относительно много веществ (гликогена, липидов), за счет расщепления которых сперматозоид частично получает энергию для движения. В хвостике при рассмотрении его в оптический микроскоп различают два отдела - главный, одетый цитоплазмой, и концевой - "голый", состоящий только из хвостовой нити.
С помощью электронного микроскопа было установлено, что тонкий чехлик, одевающий ядро сперматозоида, представляет собой уплощенную цистерну. Внутренняя ее мембрана прилежит к ядру, а наружная к плазмолемме. В задней части головки сперматозоида плазмолемма непосредственно покрывает ядро. Само ядро очень плотно заполнено переплетающимися нитями толщиной 40 Å, которые представляют собой молекулы ДНП (нуклеогистона). Химический анализ конденсированного хроматина ядра показывает, что он состоит примерно наполовину из ДНК и наполовину - из протеина. Лежащие в области шейки центриолы имеют типичное для этих органоидов строение. Меньшая из них, прилежащая сзади к ядру в его средней части, имеет вид цилиндра, образованного 9 парами трубочек. Дистальная центриола развита сильнее. Она, подобно кинетохору, находится в связи с 9 парами краев филаментов жгутика, который содержит еще 2 центральных, как и вообще все реснички и жгутики.
Плотнолежащие в промежуточном отделе вокруг осевой нити митохондрии образуют приблизительно 14 витков спирали. По-видимому, в первую очередь они обеспечивают энергией сократительные элементы сперматозоида. В главном отделе хвостика 9 пар осевых филаментов окружены кольцевыми фибриллами, которые скрепляются двумя продольными толстыми тяжами. В концевом отделе хвостика кольцевые фибриллы отсутствуют, пучок осевых фибрилл, образующих осевую нить, заключен в гомогенную массу и одет с поверхности плазмолеммой. Таким образом, здесь мы встречаемся со строением, характерным для всех ресничек и жгутиков.
Общая длина сперматозоида у человека достигает приблизительно 60 мк. Движется он активно со скоростью 3,5 мм в минуту. При этом он вращается вокруг своей оси по часовой стрелке, производя один полный оборот за 15 минут. Его способность к движению зависит от pH и других свойств среды. Продолжительность жизни сперматозоидов во влагалище составляет всего 1 час, в других женских половых путях она измеряется несколькими днями. По женским половым путям сперматозоид движется пассивно (в результате мышечных сокращений стенок матки и яйцеводов), достигая верхней трети яйцеводов, где происходит оплодотворение. Таким образом, ему не приходится тратить энергии, из ее вообще незначительного запаса, на преодоление такого длинного для него пути.
Зрелая яйцевая клетка имеет шарообразную форму; ее диаметр у человека 135 мк. Она всегда одета микроскопически видимой оболочкой и по своей структуре отличается от соматических клеток, главным образом, двумя признаками. Во-первых, в ней более или менее резко сдвинуто в пользу цитоплазмы ядерно-цитоплазменное отношение, что объясняется накоплением в теле ооцита в период роста питательных веществ для нужд будущего эмбриона. Во-вторых, в ней отсутствует клеточный центр, который исчезает в тот же период роста. Клеточный центр вносит в яйцеклетку при оплодотворении сперматозоид, и вслед за этим начинается митотическое деление зиготы - организма, находящегося на одноклеточной стадии онтогенеза.
Коротко коснемся вопроса определения пола, возникающего после оплодотворения организма. Как выяснилось, это происходит в момент зачатия, то есть оплодотворения, и обуславливается сочетанием гетерохромосом в зиготе. Обратимся к схеме (рис. 26) и рассмотрим хромосомные наборы в развивающихся женских и мужских половых клетках. Во всех оогониях имеется по две X-хромосомы. Отсюда следует, что в гаплоидный набор хромосом каждой зрелой яйцеклетки обязательно попадет одна из X-хромосом. В сперматогониях же в числе двух гетерохромосом имеется одна X-хромосома и одна Y-хромосома. Поэтому сперматозоиды должны быть разными - половина с X-хромосомой и половина с Y-хромосомой.
У млекопитающих и человека при сочетании в зиготе двух X-хромосом развивается организм женского пола, при наличии XY-хромосом - мужского. Таким образом, пол будущего ребенка будет зависеть от того, с каким из сперматозоидов отца соединится яйцеклетка матери (рис. 28). В гаплоидном наборе человека имеется та или иная из гетерохромосом (половых хромосом - гоносом) и 22 аутосомы (неполовые хромосомы). В зиготе, эмбриональных, соматических и незрелых половых клетках челолека должно быть 46 хромосом - 44+XY в мужском организме и 44+XX в женском. Лишь после первого деления мейоза в созревающих половых клетках количество хромосом редуцируется до 23. Для осуществления нормального развития необходимую наличие диплоидного набора хромосом без всяких его отклонений как количественных, так и структурных.
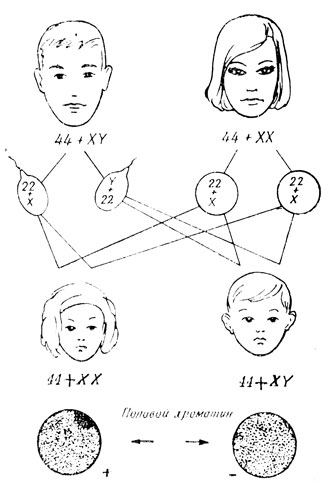
Рис. 28. Схема определения пола у человека. Половой хроматин у женщин
В 1949 году Барр установил, что у самок млекопитающих и женщин X-хромосомы в диплоидном наборе ведут себя по-разному - одна из них после деления клетки деспирализуется, подобно аутосомам, и становится неразличимой в интерфазном ядре, другая остается сильно спирализованной, подобно гетерохроматиновым хромомерам. Эта гетерохроматиновая X-хромосома, отчетливо обнаруживаемая в ядре в виде темного тельца при обычной окраске препаратов, получила название полового хроматина. В генетических опытах для выявления телец Барра (телец полового хроматина) проще всего исследовать лимфоциты крови или слущенные эпителиальные клетки. Было установлено, что любая из двух X-хромосом может стать неактивной.
В редких случаях в процессе оогенеза X-хромосомы при мейозе не расходятся. В результате этого могут образоваться яйцеклетки, отклоняющиеся от нормы: вместо одной X-хромосомы их может быть две или X-хромосома вообще отсутствует. В последнем случае при оплодотворении могут возникнуть два вида зигот, содержащих в диплоидном наборе либо одну Y-хромосому, либо одну X-хромосому, внесенную в яйцеклетку при оплодотворении сперматозоидом. Зигота с одной У-хромосомой вообще нежизнеспособна и погибает. Зигота с одной X-хромосомой у человека обладает 45 хромосомами: 44+XO. При таком сочетании хромосом из зиготы развивается неполноценная женщина малого роста с рудиментарными яичниками и, как следствие, - отсутствием вторичных половых признаков. Подобная патология известна под названием синдрома Тернера. Единственная в этом случае X-хромосома деспирализуется, и поэтому в соматических клетках половой хроматин у таких девушек не обнаруживается.
Результатом нерасхождения X-хромосом при мейозе будут и отклонения в наборе обратного характера, а именно наличие в зиготе трех X-хромосом или двух X-хромосом и одной Y-хромосомы. Женский организм с тремя X-хромосомами обозначают как "сверхсамку" или "сверхженщину" (для человека). Однако "сверхженщиной" такой индивидуум называется лишь условно, исходя из сверхкомплектного количества X-хромосом. В действительности же, при наборе хромосом 44+XXX наблюдается недоразвитие яичников и поэтому часто потеря плодовитости. Интересно, что "сверхженщины" с четырьмя X-хромосомами (хромосомный набор 44+XXXX) плодовиты, но отличаются пониженным умственным развитием. При анализе на половой хроматин соматических клеток у женщин с XXX или XXXX-хромосомами обнаруживается в ядрах соответственно 2 либо 3 тельца Барра. Таким образом, деспирализованной и активной в интерфазных ядрах и в этих случаях оказывается лишь одна из всех X-хромосом.
Зиготы с хромосомным набором типа 44+XXY развиваются у мужчин, страдающих синдромом Клайнфельтера - умственной отсталостью и недоразвитием семенника, ведущим к бесплодию. Их соматические клетки обладают половым хроматином, содержат 1 тельце Барра. Вторая X, как и у нормальных мужчин, деспирализована в интерфазе. Аналогичные дефекты развития наблюдаются и при большем числе X-хромосом, сочетающихся с Y-хромосомой, а именно при типах XXXY, XXXXY и XXXXXY. Присутствие Y-хромосомы в наборе определяет развитие особи мужского пола, но неполноценной. Количество телец Барра в интерфазных ядрах соматических клеток на единицу меньше числа X-хромосом в наборе.
Сравнительно недавно обнаружена была еще одна аномалия хромосомного набора - тип XYY. Животные "сверхсамцы" с лишней Y-хромосомой отличаются большой силой и агрессивностью. Мужчины с двумя Y-хромосомами отличаются высоким ростом (выше 180 см), большой физической силой, но пониженными умственными способностями. Как и у нормальных мужчин, интерфазные ядра их соматических клеток не содержат полового хроматина.
Учеными установлено, что передача по наследству некоторых заболеваний связана с генами, расположенными в половых хромосомах. Так, например, врожденные расстройства цветного зрения (болезнь, называвшаяся раньше дальтонизмом), вообще слепота, обусловленная атрофией зрительного нерва, гемофилия (патологическая трудноостанавливаемость кровотечений) передаются через X-хромосому.
Явление, когда в наборе имеются лишние хромосомы или какие-то из хромосом отсутствуют, носит название анеуплоидии. Наличие одной лишней хромосомы обозначают как трисомию, если таких лишних хромосом две, то это двойная трисомия. При отсутствии одной хромосомы говорят о моносомии.
Примером нарушений, связанных с увеличением в наборе числа аутосом, может служить наиболее известная трисомия - по одной из самых мелких хромосом - 21-й. Наличие в хромосомном наборе вместо двух трех 21-х хромосом имеет место при синдроме Дауна - одной из форм умственной отсталости, сочетающейся с задержкой и нарушениями физического развития, а иногда и с наличием тех или иных уродств (внешний облик больных очень сходен и характеризуется малым черепом, плоским затылком, косым разрезом глаз, широкой запавшей переносицей, полуоткрытым ртом, деформированными ушами). Половые органы недоразвиты, слабо выражены вторичные половые признаки. Половина детей с синдромом Дауна не доживает до 2 лет.
Обнаружено много заболеваний, вызываемых генами, расположенными в разных аутосомах. Среди них могут быть названы наследуемые психические болезни, такие, как шизофрения и эпилепсия.
Всевозможные наследственные заболевания человека, вызываемые различными нарушениями генотипа, являются предметом изучения усиленно разрабатываемой в настоящее время ветви цитогенетики - медицинской генетики.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'