Глава IV. Физиология клетки
Жизнь - это непрерывно текущий обмен веществ, причем процессы ассимилядии, созидания теснейшим образом связаны с диссимиляцией, распадом. Именно за счет энергии, освобождающейся при расщеплении одних веществ, строятся, синтезируются другие. Это относится в первую очередь к клеткам как элементарным живым системам, способным к самоподдержанию и воспроизведению себе подобных. Клеткам свойственны и все другие особенности, характеризующие живое - раздражимость, движение, рост и развитие. Но вся жизнедеятельность клеток в организме лишь относительно автономна и в решающей степени подчинена его нуждам и им регулируется. Та ветвь цитологии, которая изучает процессы жизнедеятельности клетки, ее функции, носит наименование цитофизиологии, или физиологии клетки.
Громадное значение для обмена веществ в клетке имеет ее проницаемость. Поступление разных химических элементов и соединений в клетку из окружающей ее среды (внутренней среды организма - кровяной плазмы либо тканевой жидкости или лимфы) и обратный их выход из клетки происходят через плазмолемму. Процесс этот очень сложный и не ограничивается простой диффузией, хотя и она имеет место - через поры плазмолеммы, из среды, где концентрация данных веществ выше, в среду с меньшей их концентрацией. Через поры плазмолеммы проходят вода и низкомолекулярные неэлектролиты.
Наряду с такой диффузией по градиенту концентрации имеет место и диффузия через плазмолемму. Таким путем могут поступать в клетки вещества, непосредственно растворяющиеся в липидных компонентах мембраны или же образующие комплексные соединения с особыми мембранными переносчиками. В силу того же концентрационного градиента образовавшееся комплексное соединение проникает в клетку. Оно тут же подвергается расщеплению на свои компоненты. И снова вследствие выравнивания концентрации мембранный переносчик возвращается на поверхность плазмолеммы, а вещество, проникшее таким способом внутрь клетки, включается в ее метаболизм.
Благодаря системе активного переноса во многих клетках постоянно поддерживается концентрация калия гораздо более высокая, чем в окружающей среде, а натрия, - наоборот, более низкая. Клетка активно поглощает ионы калия и активно выводит в окружающую среду ионы натрия. Это явление получило образное название калиевого и натриевого насоса.
Мембрана, одевающая клетку - ее плазмолемма, избирательно пропускает через себя, причем в обе стороны, воду и многие растворенные в ней вещества. Благодаря этому в клетке быстро ликвидируются нарушения оптимального для нее осмотического давления.
Поступление веществ в клетку из окружающей среды определяется в большой мере степенью их связывания (путем химического соединения или адсорбции) с протоплазмой, а также быстротой утилизации в процессе метаболизма.
Обменные процессы, которые протекают в клетке за счет веществ, поступающих в нее извне, носят двоякий характер. Это, во-первых, реакции расщепления сложных веществ на простые. Такие реакции обычно протекают с освобождением энергии, которая необходима клетке для биохимических процессов обратного характера, созидательных. Непрерывно текущие в живой клетке реакции расщепления - это форма метаболизма, которую обозначают как ее энергетический обмен. Другая форма клеточного метаболизма - пластический обмен. К нему относятся все виды биосинтетических процессов, в частности, связанных с восстановлением самой протоплазмы. Пластический обмен осуществляется за счет энергетического, причем для биохимического синтеза широко используются те низкомолекулярные соединения, которые возникают при расщеплении высокомолекулярных: аминокислоты - при расщеплении белков, нуклеотиды - при расщеплении нуклеиновых кислот и т. п. Из этих аминокислот и нуклеотидов строятся соответственно новые белковые молекулы и новые цепи ДНК и РНК, специфичные по своей структуре для клеток данного организма. Раскрытие процесса белкового синтеза клетками явилось величайшим достижением современной биологии и открыло новый, решающий этап в ее развитии как науки о живом.
В процессе эволюции живых существ на Земле сложился, как выяснилось в последние годы, особый аппарат синтеза белка, в принципе единый не только у одноклеточных и многоклеточных организмов, но и у живых существ с доклеточной организацией тела - бактерий, вирусов и фагов.
Синтез белка происходит с помощью нуклеиновых кислот, из которых молекулы ДНК являются носителями наследственных свойств, присущих данному виду организмов, а молекулы РНК - подсобными элементами, с помощью которых ДНК клетки осуществляет свою функцию. Как выяснилось, разное расположение четырех нуклеотидов в молекуле ДНК представляет собой шифр, код, с помощью которого зашифрована, записана, как теперь принято говорить, наследственная информация, т. е. наследственные свойства организма и каждой его клетки в отдельности. Синтез того или иного белка кодируется расположением аденина (А), гуанина (Г), тимина (Т) и цитозина (Ц) в данном участке двунитчатой спирали молекулы ДНК.
Никого не удивляет, что с помощью алфавита из 32 букв можно сделать бесконечное множество описаний. При помощи всего двух знаков азбуки Морзе (тире и точки) можно передать любой текст. Оперируя только двумя знаками (0 и 1), можно заложить в электронносчетную машину неограниченную информацию. Поэтому совсем не является парадоксальной обнаруженная биохимиками и молекулярными биологами в живой природе запись наследственной информации в молекулах ДНК с помощью четырех знаков. Причем было установлено, что (каждый раз для кодирования той или иной аминокислоты используются всего 3 из указанных выше азотистых оснований, т. е. триплеты их.
Однако оказалось, что дело обстоит далеко не так просто: кодирует расположение аминокислот в цепи белковой молекулы не сама ДНК, а молекулы РНК, которые строятся молекулой ДНК- Нити молекулы ДНК ведут себя по-разному. Молекулы РНК строятся только вдоль одной из этих нитей, а вторая используется лишь в процессе редупликации самой ДНК. Молекулы РНК, в отличие от молекул ДНК, являются одноцепочечными. Из образующих их нуклеотидов четырех видов три являются общими с ДНК - А, Г и Ц, и только вместо Т здесь имеется азотистое основание урацил (У). При синтезе молекулой ДНК молекулы РНК против молекулы А становится У, а против молекулы Г и Ц соответственно Ц или Г. Участок новообразованной молекулы РНК отвечает комплементарно (дополнительно) по составу нуклеотидов синтезировавшему ее участку одной из цепочек развернувшейся молекулы ДНК.
Молекулы РНК несравненно короче молекул ДНК, причем было установлено, что разным из них принадлежит различная функция в процессе синтеза белка. Более длинные из них обладают молекулярным весом, равным нескольким сотням тысяч. Это так называемая матричная, или информационная РНК, обозначаемая как и-РНК. Кроме того, имеется РНК - переносчик аминокислот, или транспортная, ее символ т-РНК. Молекулы последней совсем незначительных размеров, и молекулярный вес их составляет приблизительно 25 000.
Молекулой ДНК при участии ядрышкового аппарата образуется и особая РНК, входящая в состав рибосом. В ядрышке к этой РНК присоединяется синтезированный им специфический рибосомный белок, т. е. формируются рибосомы. Функцией ядрышка клетки является, таким образом, как биосинтез обоих компонентов рибосом, так и сборка их самих. Недавно было обнаружено, что у тех редких объектов, где ядрышки отсутствуют, рибосомы вообще не образуются.
Рибосомы, которые некоторое время входят в состав ядрышка, затем выходят из него в кариолимфу, а отсюда поступают через поры в ядерной оболочке в окружающую цитоплазму и распределяются по ней, частично оседая на наружной поверхности канальцев и цистерн шероховатого эндоплазматического ретикулума и на наружной ядерной мембране.
Рибосомы - самые тяжелые из молекул РНК. Молекулярный вес их доходит до полутора миллионов. Функционируют рибосомы не в одиночку, а группами, которые получили название полисом. Молекула и-РНК, образуемая молекулой ДНК (рис. 24А), связывается с полисомой и здесь начинает свою деятельность, которая протекает с помощью молекул т-РНК. Количество видов транспортных РНК, или РНК-переносчиков, соответствует числу 20 известных аминокислот. Каждая из т-РНК присоединяет к себе молекулу активированной ферментами аминокислоты соответствующего ей вида, а затем несет эту аминокислоту к рибосоме, где укладывает ее вдоль нити информационной РНК, прилежащей к полисоме. Каждая из рибосом последовательно перемещается вдоль молекулы информационной РНК по мере декодирования, "считывания" с последней содержащейся в ней информации. Информационная РНК может служить матрицей для синтеза белка несколько раз, после чего она разрушается. Все эти сложные явления, разыгрывающиеся в строгой последовательности на пути образования живым веществом клетки белковых молекул, осуществляются за счет энергии молекул как АТФ, так и другого нуклеозидфосфата - ГТФ (гуанозинтрифосфорной кислоты), синтезируемых митохондриями.
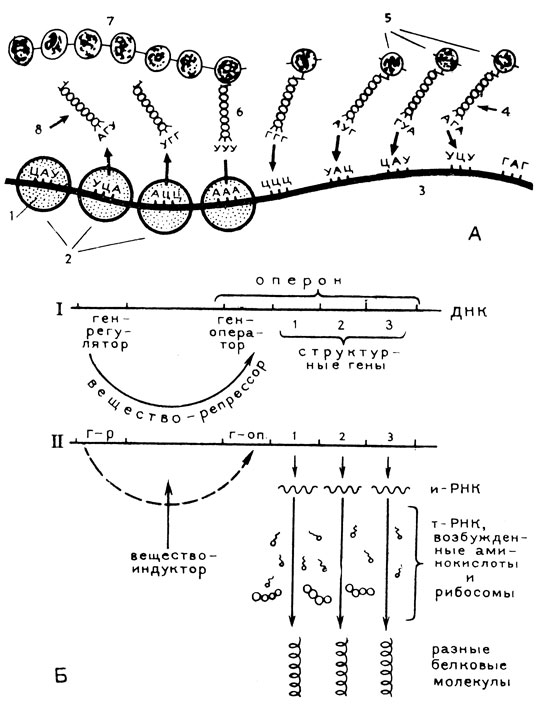
Рис. 24. А - схема синтеза белка: 1 - рибосома; 2 - полисома; 3 - молекула и-РНК; 4 - молекула т-РНК, присоединившая к себе молекулу активированной ферментами аминокислоты (5); 6 - молекулы т-РНК, присоединившиеся к комплементарным участкам молекулы и-РНК; 7 - растущая полипептидная (белковая) цепочка из соединившихся друг с другом аминокислот; 8 - молекула т-РНК, освободившаяся от аминокислоты и молекулы и-РНК. Б - схема регуляции синтеза белка по Жакобу и Моно: I - синтез заблокирован, II - синтез в действии
Молекула транспортной РНК содержит в своей относительно короткой нити те же нуклеотиды, что и молекула информационной РНК: А, У, Г и Ц. Цепочка т-РНК сложена таким образом, что с одного ее конца располагается участок, несущий аминокислоту, а с другого - участок, определяющий "сродство" данной т-РНК определенному триплету и-РНК, иначе говоря, комплементарный этому триплету. Подходя к цепочке и-РНК, молекула т-РНК связывается с ней своим триплетом таким образом, что против А в триплете и-РНК ложится У триплета т-РНК, а против Ц ложится Г и т. д. Оказавшиеся рядом в этом конвейере аминокислоты соединяются друг с другом. Далее в соответствии с расположением вдоль молекулы и-РНК тех или иных азотистых оснований последовательно присоединяются к ней новые коротенькие молекулы РНК, транспортирующие соответствующие им аминокислоты, которые в свою очередь устанавливаются на свое место в строящуюся белковую цепочку.
Таким образом, состав белковой молекулы и последовательность расположения в ней аминокислот диктуются порядком расположения в молекуле информационной РНК азотистых оснований, точнее их триплетов. Так, если искусственно создать в лаборатории нить РНК, в которой против остатков фосфорной кислоты будут стоять только урацилы, строящаяся в этих условиях белковая молекула (вернее, полипептидная цепочка) будет состоять только из аминокислоты фенилаланина. Таким путем и было установлено, что триплет, кодирующий аминокислоту фенилаланин, т. е. определяющий ее место в синтезируемой белковой молекуле, состоит из трех рядом стоящих урацилов, что символически изображается как УУУ. Путем соответствующих экспериментов были расшифрованы и другие триплеты для всех 20 аминокислот. При этом оказалось, что некоторые из аминокислот кодируются не одним триплетом, что увеличивает надежность этого процесса. Так, например, аминокислота лизин кодируется триплетами ААА или ААГ, глютаминовая кислота - триплетами ГАА или ГАГ. К этим кодонам (кодирующим триплетам нуклеотидов) и-РНК пристраиваются комплементарные, т. е. соответствующие им антикодоны транспортных РНК. В случае лизина это т-РНК, несущие УУУ или УУЦ, а в случае глютаминовой кислоты - т-РНК, несущие ЦУУ или ЦУЦ.
Нить молекулы ДНК на разных участках длины строит разные (по последовательности расположения в них азотистых оснований) молекулы и-РНК. Таким образом, молекула ДНК может управлять синтезом большого количества разных белков, а поскольку комбинации в последовательности расположения нуклеотидов в молекулах ДНК могут быть исключительно разнообразными, то это, в результате использования через соответствующих посредников молекул и-РНК, приводит к такому же разнообразию расположения аминокислот в белковой молекуле, т. е. фактически к безграничному разнообразию носителей жизненных свойств - белков.
Процесс "считывания" с ДНК на РНК называют транскрипцией. Представление о том, что ДНК определяет синтез РНК, а РНК далее определяет синтез белка, стало основным положением современной молекулярной биологии.
Сопоставление соматических клеток в ряду животных показывает, что в процессе филогенеза, исторического развития от губки до позвоночных включительно шло непрерывное увеличение содержания ДНК в клетке, хотя код ДНК особых изменений при этом не претерпел. Одним из факторов, определяющих количество ДНК, содержащегося в ядре, является, как выяснилось, количество цитоплазмы, находящейся под контролем ядра.
Как было установлено, очень длинная нить развернувшейся молекулы ДНК, на которой строятся молекулы информационной РНК, одновременно работает только отдельными своими участками. Согласно широко принятым в настоящее время представлениям, высказанным в 1961 году французскими учеными Жакобом и Моно, в указанной нити ДНК чередуются отрезки, обозначаемые как опероны и гены-регуляторы. Каждый оперон в свою очередь состоит из гена-оператора и следующих за ним структурных генов (рис. 24Б), которых может быть разное число. Структурный ген - это именно тот участок нити ДНК, где строится информационная РНК, по шаблону которой в дальнейшем на полисоме собирается молекула определенного белка. В обычном состоянии структурные гены не работают, так как по гипотезе Жакоба и Моно ген-регулятор вырабатывает особое вещество - репрессор, воздействующий как тормоз на ген-оператор, а включение структурных генов к действию происходит лишь по сигналу из гена-оператора, который командует таким образом всеми структурными генами оперона. Связь между геном-регулятором и геном-оператором нарушается лишь при появлении в среде вещества-индуктора. В результате взаимодействия индуктора и репрессора возникает продукт, уже лишенный репрессорной активности. После снятия репрессора оператор начинает оказывать активизирующее влияние на весь ряд контролируемых им структурных генов. Обычно ряд этот вырабатывает молекулы белков, последовательно входящих в одну какую-то реакцию.
В недавнее время было сделано очень важное открытие - обнаружена возможность передачи в клетке информации обратного характера - с РНК на ДНК. Синтез молекулы ДНК (гена) на молекуле РНК осуществляется с помощью фермента, названного обратной транскриптазой. Некоторые крупные советские ученые (Н. П. Дубинин, А. Н. Студитский) видят принципиальное значение обратного синтеза ДНК с и-РНК в том, что, по их мнению, и-РНК, являющиеся рабочими молекулами в клетке, способны изменяться во время работы. Изменение же молекулы и-РНК, строящей генную молекулу, должно приводить к изменению этого гена, т. е. к изменению наследственной информации, записанной в молекуле ДНК. Однако возможность упомянутых функционально обусловленных изменений структуры молекул и-РНК еще требует доказательств.
Мы уже отмечали, что ферменты - это белки. В клетках обнаружены тысячи различных ферментов, которые являются активнейшими катализаторами биохимических процессов. Благодаря ферментам различного рода реакции в клетке протекают с чрезвычайной быстротой, мгновенно, так что для образности иногда говорят о "вихре" клеточного обмена веществ. Как пластический, так и энергетический обмен в клетке осуществляются поэтапно, и переход от одного звена к другому катализируется особым ферментом. Соответственно этому каждый фермент вырабатывается отдельным структурным геном, а вся цепь ферментов, обеспечивающих протекание какого-то целостного биохимического процесса, синтезируется последовательно единым опероном.
В последнее время многие исследователи на основании экспериментов высказывают точку зрения, что репрессорами генов являются белки гистоны, входящие в состав хромосом. Эти белки были открыты около 100 лет тому назад Мишером.
В настоящее время установлено, что все клетки каждого организма, за редким исключением, содержат в общем одинаковые типы гистонов. Различаются они лишь количественным соотношением гистона и ДНК. С другой стороны, оказалось, что и разные организмы обладают сходными гистонами. Интересно, что синтез гистонов происходит в клетке во время интерфазы в ядрышке и протекает синхронно с синтезом ДНК, причем новообразованная молекула ДНК покрывается как бы оболочкой из гистона.
Таким образом, поскольку сейчас, согласно мнению многих молекулярных биологов, активность различных участков ДНК-оперонов трактуется как находящаяся в зависимости от гистонов, на очередь ставится вопрос о том, каким же образом осуществляется это взаимодействие в нуклеогистоне между его компонентами.
Известный советский цитолог В. Я. Бродский (1966) на основании собственных экспериментов и данных литературы пришел к выводу, что имеются клетки с преимущественно цитоплазменным или же с преимущественно ядерным типом синтеза белка. Он предложил схему некоторых морфологических особенностей клеток с преимущественно той или иной локализацией белкового синтеза. Автор перечисляет следующие основные особенности большинства клеток с преимущественно ядерным типом синтеза: 1. Преобладание размеров и массы ядра над размерами цитоплазматической части клетки. 2. Увеличение поверхности ядра за счет выростов и выпячиваний ядерной оболочки как микроскопических, так и субмикроскопических. 3. Усиление контактов ядерной оболочки с цитоплазматическими структурами, в частности, с митохондриями (большинство описанных в литературе контактов оболочки ядра с митохондриями относится именно к клеткам с преимущественно ядерной локализацией синтеза белка - эмбриональным, дифференцирующимся, опухолевым). 4. Высокая концентрация РНК и свободных нуклеотидов в ядре. Значительное содержание в нем субмикроскопических гранул. 5. Относительно слабое развитие эндоплазматической сети, диффузное распределение рибосом в гиалоплазме. Рибосомы, как правило, не связаны с мембранными структурами. Полисомы встречаются сравнительно редко. 6. Низкая суммарная выработка белка сравнительно с клетками цитоплазматического типа.
Клетки с цитоплазматическим типом синтеза белка отличаются, наоборот, развитой эргастоплазмой (густым гранулярным эндоплазматическим ретикулумом) и значительным преобладанием РНК в цитоплазме по сравнению с ядром. По образному выражению автора, это настоящие фабрики по производству белка. Прежде всего это железистые клетки, образующие белковый секрет (например, поджелудочной железы). Дифференцированные клетки с цитоплазматическим типом синтеза белка образуют его в большинстве случаев не для своих собственных нужд, а в интересах целого организма. Этот белок (например, секрет или антитела) затем выводится из клетки. Особое место в этом отношении занимают такие большие клетки, как нервные. Усиленный синтез белка в них обусловлен необходимостью непрерывно восстанавливать снашивающиеся в процессе жизнедеятельности большие массы белка их крупных отростков. Как клетки с преимущественным синтезом белка должны быть также упомянуты ооциты.
В. Я. Бродский подчеркивает, что в процессе клеточной дифференцировки соотношение ядерной или цитоплазматической локализации синтеза белка может измениться. В качестве примера он приводит эмбриональные клетки с преимущественно ядерным типом белкового синтеза, в которых по мере дифференцировки и специализации основной синтез белка переходит к цитоплазме. Другой пример - плазматические клетки (рис. 13), синтезирующие большое количество белков - антител. Они имеют строение, характерное для клеток с цитоплазматическим типом синтеза белка. Их же предшественники - первичные плазмобласты - являются клетками с типично ядерной локализацией белкового синтеза.
Показателем усиливающейся продукции белка клеткой может служить увеличение концентрации РНК в месте его синтеза.
Говоря о функции пластинчатого комплекса, мы уже отмечали его роль как органоида клетки, в котором происходит в основном "сборка" разного рода веществ, вырабатываемых клеткой. В качестве одного из примеров мы приводили оформление в нем гранул белкового секрета. Очень наглядно показано это на схеме одного из крупнейших современных цитологов Гирша (1959). Согласно данной схеме вещества, из которых строится секрет, клетка получает из крови или из непосредственно омывающей ее тканевой лимфы. За счет этих материалов и энергии макроэргических молекул, синтезируемых митохондриями, на рибосомах гранулярного ретикулума и на полисомах строятся белковые молекулы. Они поступают в канальцы эргастоплазмы, где накапливаются в форме первичного секрета. Увеличивающиеся благодаря этому в диаметре канальцы гранулярного эндоплазматического ретикулума расширяются и принимают вид цистерн. По схеме Гирша цистерны образуют местные разрывы, и первичный секрет непосредственно поступает в цистерны пластинчатого комплекса. По другим данным первичный белковый секрет из канальцев гранулярного эндоплазматического ретикулума поступает в канальцы агранулярного и лишь оттуда в цистерны пластинчатого комплекса. Здесь первичный секрет превращается в относительно плотные гранулы вторичного белкового секрета путем постепенного укрупнения, сопровождающегося одновременно постепенной значительной потерей воды. Гранулы покидают пластинчатый комплекс, одетые липопротеиновой мембраной. В процессе выхода такой гранулы из клетки ее липопротеиновая мембрана первоначально сливается с плазмолеммой в месте их контакта. Затем происходит небольшой разрыв плазмолеммы, и секрет выводится наружу.
Синтез жирового секрета осуществляется в канальцах и цистернах агранулярного эндоплазматического ретикулума. Фосфолипиды, как и полисахариды, синтезируются самим пластинчатым комплексом.
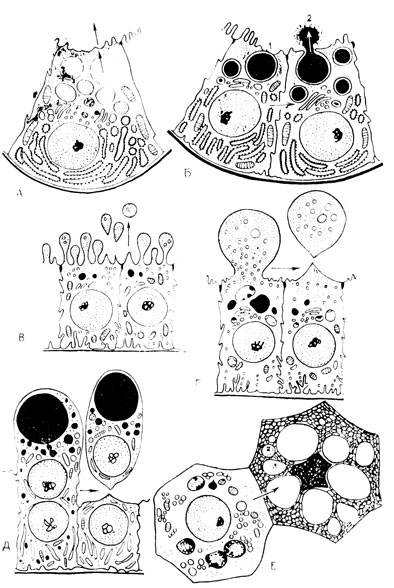
Рис. 25. Схема (в основном по Курасуми) различных типов секреторного процесса в железистой клетке, по данным электронной микроскопии: А и Б - мерокринная (эккринная) секреция, В, Г и Д - апокринная секреция, Е - голокринная секреция
Выведение секрета из клетки может происходить как с разрывом плазмолеммы, так и без него (рис. 25). В последнем случае образовавшийся вышеописанным способом белковый секрет проходит через плазмолемму, не разрушая ее, что осуществляется уже на молекулярном уровне. Оба эти способа получили наименование мерокринной (по-гречески "мерос" - часть, "крино" - выделяю) секреции. Выведение синтезированных клеткой продуктов может быть связано с отрывом выпячиваний цитоплазмы, где они скапливаются. Такой способ выведения секрета носит название апокринного ("апокс" - верхушка).
Различают секрецию микро- и макроапокринную. При микроапокринной секреции отрыв ограничивается разбухшими концами микроворсинок. Макроапокринная секреция осуществляется отрывом всего "наплыва" цитоплазмы, которая таким образом сама принимает непосредственное участие в образовании конечного секрета. В некоторых случаях (в лактирующей молочной) может отрываться значительная, даже большая часть клетки вместе с находящимися здесь органоидами - митохондриями и элементами пластинчатого комплекса.
Все они вместе с синтезированным клеткой белковым и жировым секретом дают начало сложному секрету молока. Наконец, встречается еще один способ секреции - голокринный (по-гречески "голос" означает весь). В этом случае происходит полная гибель клеточного тела и превращение его в секрет, например, в капли жира в сальных железах.
В клетках, секретирующих по мерокринному типу, могут наблюдаться специальные структуры, способствующие выведению секрета. Такая система внутриклеточных капилляров имеется, например, в так называемых обкладочных клетках трубчатых желез дна желудка.
Секретировать может и ядро клетки. Было описано, в частности, образование жировых капель в ядрах клеток лактирующей молочной железы собаки.
Раздражимость - это свойство живого, в частности клеток, воспринимать воздействия изменяющейся окружающей среды и реагировать на них. Раздражителями являются факторы внешней для клетки среды, вызывающие ее реакцию. Степень чувствительности восприятия раздражений и формы реакции зависят от многих причин, в том числе от видовой, и особенно тканевой принадлежности клеток, а также от их физиологического состояния.
В процессе исторического развития самостоятельно живущие клетки - клетки-организмы дали начало многоклеточным животным. Дальнейшее филогенетическое становление тканей, как функциональных систем клеток, шло на основе разделения функций между клетками и их интенсификации. Произошла специализация клеток, отражающая их тканевую природу.
Реакции клеток на внешние раздражения носят специфический характер и строго соответствуют их морфофункциональной специализации. Однако помимо этого клетки самых различных тканей выявляют в состоянии возбуждения общие для всех них субстратные изменения. Поверхность клетки приобретает отрицательный электрический заряд вместо положительного. Наблюдается укрупнение белковых частиц протоплазмы в результате изменения их электрических зарядов, увеличивается клеточная проницаемость и т. д. При большой силе раздражающего воздействия наступает повреждение клетки и все признаки, характеризующие ее возбужденное состояние, резко усиливаются. Понижение дисперсности протоплазмы достигает такой степени, что она мутнеет. Происходит повышение вязкости и сорбционных свойств живого вещества клетки, реакция ее становится кислой в результате нарушения обмена веществ и накопления молочной кислоты. Однако состояние это обратимо, и после устранения действия раздражителя клетка возвращается по всем показателям к норме. Д. Н. Насонов и его ближайший ученик и сотрудник В. Я. Александров, много занимавшиеся изучением описанного состояния клетки, назвали его паранекрозом (по-гречески "пара" - около, "некрос" - смерть). Они выдвинули денатурационную теорию повреждения, согласно которой повреждение клетки представляет собой обратимую денатурацию ее белков.
Белки являются полимерами. Их первичная структура представлена длинной цепью мономеров - аминокислот, которые прочно соединены друг с другом. Эта цепь закручена в спираль, что составляет вторичную структуру белка. Наконец, спираль свертывается в шарик-глобулу. В такой уже третичной структуре расположение спиралей специфично для каждого белка. Во вторичной и третичной структурах в отличие от первичной химические связи слабые, и они сравнительно легко рвутся при различных воздействиях на клетку. Тогда глобула развертывается и раскручивается - происходит денатурация белка.
О состоянии паранекроза очень удобно судить, применив прижизненную окраску, например, нейтральным красным. Если клетка находится в нормальном состоянии, проникающий в нее витальный краситель она отмешивает в виде вакуолей и гранул, которые накапливаются в цитоплазме. Цитоплазма и ядро клетки благодаря этому остаются неокрашенными. В состоянии паранекроза этот механизм, очень важный для благополучия клетки, оказывается выключенным. Клетка лишается способности самоочищения от проникающего в нее красителя - гранулы отмешанной краски не образуются, цитоплазма и ядро окрашиваются диффузно. При этом ядра выявляют структуру, характерную для фиксированного препарата, свидетельствующую о резком снижении дисперсности кариоплазмы. В отличие от фиксированного состояния после устранения агента, вызвавшего паранекроз, цитоплазма и ядро раскрашиваются, и вся краска, проникшая в клетку, отмешивается в гранулы. Ядро снова становится оптически пустым.
Основываясь на общности картин, наблюдающихся при возбуждении и повреждении клеток, Д. Н. Насонов и В. Я. Александров пришли к заключению, что и возбуждение клеток обусловлено обратимой денатурацией ее белков. Денатурационная теория повреждения и возбуждения приобрела в настоящее время многих последователей как в Советском Союзе, так и за рубежом.
Наиболее примитивной формой клеточного движения является амебоидное. Оно состоит в постепенном перетекании клеточного тела по направлению образуемых цитоплазмой выпячиваний - ложноножек-псевдоподий. Такой тип движения присущ белым клеткам крови и блуждающим клеткам соединительной ткани.
Скользящее движение обнаружено у оседлых волокнообразующих клеток соединительной ткани - фибробластов. Как оно происходит, пока не установлено.
Движение с помощью жгутика у млекопитающих и человека присуще только мужским половым клеткам - сперматозоидам.
Другие виды клеточного движения не связаны с перемещением клетки в пространстве. Они осуществляются клетками в интересах целого организма. Это ресничное движение и мышечное сокращение.
Что же касается внутриклеточного движения, то со многими формами его мы уже познакомились раньше. Это движение митохондриев по клетке, перемещение митотического аппарата и хромосом во время митоза, раскручивание молекулы ДНК при ее редупликации и построении молекул разных РНК, движение всех видов РНК из ядра и по цитоплазме в процессе синтеза белка, движение белковых молекул при их обратимой денатурации и т. д.
К росту способны все клетки. Дочерние клетки после митоза, в период интерфазы, вырастают до размеров материнской. Именно до этой величины, но не больше. В конце прошлого века Дриш (1899) выдвинул положение о постоянстве размеров клеток. Это касается, конечно, клеток конкретных тканей животных соответствующего вида. То же самое наблюдается и у людей. Если измерить слущенные клетки ротовой полости человека 2-метрового роста и 1-метрового карлика, они окажутся одинаковой величины. Изучение полиплоидных клеток показало, что с удвоением плоидности происходит и увеличение их объема. В результате пришлось переформулировать правило Дриша о постоянстве клеточных размеров и говорить о постоянстве минимальных клеточных размеров (Е. М. Вермель, 1934). Имеются в виду, конечно, зрелые клетки.
С увеличением объема клетки соотношение между массой протоплазмы и клеточной поверхностью изменяется, так как объем шара, как известно, увеличивается пропорционально кубу радиуса, а поверхность его - пропорционально квадрату радиуса. Поверхность, таким образом, отстает от объема и, следовательно, затрудняются обменные процессы между клеткой и окружающей ее средой. Были высказаны соображения, что возникающие затруднения в указанных обменных взаимоотношениях и являются стимулом к митозу у клеток, достигших предельных благоприятных размеров.
Однако клетки могут в известных пределах изменять свои размеры в зависимости от функционального состояния, например, увеличиваться при усилении их деятельности. В этом случае изменяется ядерно-цитоплазменное отношение в пользу ядра. Сильнее всего увеличено ядрышко, в его пользу сдвигается ядрышково-ядерное отношение.
С другой стороны, имеются "летки, рост которых увеличивается по мере роста организма. Это нервные клетки, уже не способные к размножению в постнатальном периоде. Увеличение размеров многих нервных клеток связано с их полиплоидизацией. Фаза синтеза в нервных клетках может длиться десятки лет. На примере нервных клеток хорошо видно, что полиплоидизация - явление, носящее компенсаторный характер. Нервные клетки, уже не способные к митотическому делению, благодаря полиплоидизации увеличивают свои размеры и рабочую производительность. Возникающие при этом затруднения в ядерно-цитоплазменном обмене могут разрешаться путем амитотического разделения ядра и возникновения таким путем дву- и многоядерности нервных клеток. Такое раздробление ядра приводит к увеличению поверхности контакта между ядром и цитоплазмой.
Увеличиваются с возрастом и размеры мышечных волокон, количество которых в скелетной мускулатуре мало изменяется после рождения. На примере волокон мышечной ткани особенно наглядно видна способность к росту благодаря усиленной функции. Такой "рабочий" рост получил название физиологической гипертрофии.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'